- Insekten
-
Insekten 
Honigbiene (Apis mellifera)
Systematik ohne Rang: Urmünder (Protostomia) Überstamm: Häutungstiere (Ecdysozoa) Stamm: Gliederfüßer (Arthropoda) Unterstamm: Tracheentiere (Tracheata) Überklasse: Sechsfüßer (Hexapoda) Klasse: Insekten Wissenschaftlicher Name Insecta (Linnaeus, 1758) Unterklassen - Felsenspringer (Archaeognatha)
- Fischchen (Zygentoma)
- Fluginsekten (Pterygota)
Insekten (Insecta), auch Kerbtiere oder Kerfen genannt, sind die artenreichste Klasse der Gliederfüßer (Arthropoda) und zugleich die mit absoluter Mehrheit auch artenreichste Klasse der Tiere überhaupt. Beinahe eine Million Insektenarten sind bisher wissenschaftlich beschrieben worden (925.000 nach[1], 865.000 nach[2]). Damit sind mehr als 60% aller beschriebenen Tierarten Insekten. Nach verschiedenen Hochrechnungen rechnet man allerdings mit einem Vielfachen, wobei vor allem in den tropischen Regenwäldern noch Millionen unentdeckter Arten vermutet werden. Fossil lassen sich Insekten zum ersten Mal vor rund 400 Millionen Jahren im Devon nachweisen.
Der Name „Insekt“ wurde im 18. Jahrhundert aus dem lateinischen insectum eingedeutscht. Dies gehört zu in-secare = „einschneiden“ und bedeutet demnach eigentlich „eingeschnittenes (Tier)“, was sich auf die stark voneinander abgesetzten Körperteile bezieht. Es ist eine Lehnübersetzung vom griechischen éntomon = „Insekt“ (zu entémnein = „einschneiden“), was sich heute in Begriffen wie der Entomologie (Insektenkunde) spiegelt. Der Begriff „Kerbtier“ geht auf den deutschen Schriftsteller Philipp von Zesen zurück. Früher wurde auch der wissenschaftliche Name Hexapoda (griechisch für „Sechsfüßer“) verwendet, der heute für eine übergeordnete Gruppe reserviert ist (siehe dazu die Anmerkung im Systematik-Abschnitt weiter unten).
Inhaltsverzeichnis
Äußere Anatomie
 Schema zur Anatomie der Insekten
Schema zur Anatomie der Insekten
A – Caput (Kopf)
B – Thorax (Brust)
C – Abdomen (Hinterleib)
1. Antenne
2. Ocellus (vorne)
3. Ocellus (oben)
4. Komplexauge (Facettenauge)
5. Gehirn
6. Prothorax
7. rückseitige (dorsale) Arterie
8. Tracheen
9. Mesothorax
10. Metathorax
11. Erstes Flügelpaar
12. Zweites Flügelpaar
13. Mitteldarm
14. Herz
15. Eierstock
16. Hinterdarm (Rektum)
17. Anus
18. Vagina
19. bauchseitiges Nervensystem mit Ganglien
20. Malpighische Gefäße
21. Tarsomer
22. Prätarsus
23. Tarsus
24. Tibia
25. Femur
26. Trochanter
27. Vorderdarm
28. Thoraxganglion
29. Coxa
30. Speicheldrüse
31. Unterschlundganglion
32. MundwerkzeugeDie Größe der Insekten variiert sehr stark und liegt bei den meisten Arten zwischen einem und 20 Millimetern. Die kleinsten bekannten Arten sind dabei Vertreter der zu den Käfern gehörenden Federflügler sowie Erzwespen mit Körperlängen um 0,2 Millimeter. Die größten bekannten Insekten sind Stabheuschrecken mit etwa 33 Zentimetern Körperlänge sowie der Bockkäfer Titanus giganteus mit einer Körperlänge von 16 Zentimeter und einer Breite von etwa 6 Zentimetern. Entsprechend ihrer Lebensweise kann der Körper der Insekten langgestreckt, abgeflacht oder mehr oder weniger kugelig sein.
Allen Insekten gemeinsam ist die meist deutlich sichtbare Gliederung des Leibes in die drei Abschnitte (Tagmata) Kopf (Caput), Brust (Thorax) und Hinterleib (Abdomen), sowie der feste Chitinpanzer und das Vorhandensein von genau drei Beinpaaren. Alle Körperteile bestehen aus einzelnen Segmenten, die eine Rückenplatte (Tergit), eine Bauchplatte (Sternit) sowie Seitenplatten (Pleurite) aufweisen. Seitlich der einzelnen Segmente sind im Grundbauplan Öffnungen des Tracheensystems, so genannte Stigmen, angelegt, deren tatsächliche Anzahl jedoch bei den einzelnen Taxa der Insekten stark schwanken kann. Auch Extremitäten und deren Teile (Mundwerkzeuge, Styli, Gonopoden) können an jedem einzelnen Segment jeweils in Paaren vorkommen. Das Brustsegment trägt sechs Beine (drei Paare). Jedes Segment besitzt außerdem ein Ganglienpaar im Bauchmark, wobei die Ganglien des Kopfes zu einem Oberschlund- und einem Unterschlundganglion verschmolzen sind. Zwischen den einzelnen Segmenten befinden sich dehnbare Häutchen, die Intersegmentalhäute, die eine Beweglichkeit der Segmente gegeneinander sowie eine Volumenveränderung des Körpers bei der Eiproduktion, Nahrungsaufnahme oder Atmung ermöglichen. Bei starr verschmolzenen Segmenten, etwa im Kopf, sind diese Häutchen nicht vorhanden.
Der Chitinpanzer, der neben Chitin auch mehrere Proteine enthält, bildet die äußere Schicht des Insektenkörpers. Er wird von einer darunter liegenden Epidermis gebildet, und in ihn eingebettet befinden sich die Sinnesorgane sowie verschiedene Drüsen des Insekts. Mit mehr oder weniger starker Beteiligung der Epidermis bildet der Chitinpanzer verschiedene Oberflächenstrukturen aus, zu denen Warzen, Dornen, Haare, Borsten, Schuppen und Höcker gehören. Durch die Einlagerung von Farbstoffen (Pigmentfarben) oder aufgrund spezieller lichtbrechender Oberflächen (Interferenzfarben) können die Außenschicht des Insekts oder einzelne Körperteile gefärbt sein.
Als Sinnesorgane dienen vor allem Haarsensillen, die über den Körper verteilt sind. Diese reagieren auf Erschütterungen und Schwingungen, können jedoch auch Gerüche, Feuchtigkeit oder Temperaturen wahrnehmen. Einige dieser Sinneszellen sind zu Sinnesorganen gruppiert, so etwa das Johnstonsche Organ am Pedicellus der Antenne oder die Tympanalorgane zur Geräuschwahrnehmung, die man beispielsweise bei den Langfühlerschrecken findet. Dadurch ist die Wahrnehmung von Schwingungen im Bereich von 1 Hz bis 100.000 Hz möglich. Als optische Sinnesorgane dienen die Facettenaugen sowie die Ocellen, bei vielen Larven auch die larvalen Punktaugen.
Der Insektenkopf: Augen, Antennen, Mundwerkzeuge
 Kopf einer Libelle mit Facettenauge und Ocellen
Kopf einer Libelle mit Facettenauge und OcellenDer Insektenkopf besteht aus einem vorderen Abschnitt, dessen Segmentnatur umstritten ist (nach der früher vorherrschenden Articulata-Hypothese entspricht er dem Acron der Anneliden) und fünf weiteren miteinander verschmolzenen Segmenten und trägt die Augen und die Gliederantenne sowie einen typischen Apparat von Mundwerkzeugen aus paarigen Mandibeln und Maxillen sowie einem unpaaren Labium[3]. Entsprechend befindet sich am Kopf auch die Mundöffnung, durch die Nahrung aufgenommen wird. Die von außen sichtbaren Nähte des Insektenkopfes haben mit den Segmentgrenzen bis auf eine Ausnahme, der Grenze zwischen dem fünften und sechsten Segment (Postoccipitalnaht), nichts zu tun.
Die Facettenaugen sitzen auf dem vorderen Abschnitt, daneben besitzen Insekten im Grundbauplan drei Punktaugen (Ocellen) zur Hell-Dunkelwahrnehmung. Das zweite Segment ist das Antennalsegment mit der für die Insekten typischen Geißelantenne. Diese unterscheidet sich von der Gliederantenne, die man bei den ursprünglichen Hexapoden findet, dadurch, dass sie nur ein Basal- oder auch Schaftglied mit Muskulatur besitzt, den Scapus. Das darauf folgende Wendeglied, der Pedicellus, ist gemeinsam mit der Geißel gegenüber dem Scapus beweglich. Im Pedicellus befindet sich das Johnstonsche Organ, ein Sinnesorgan, welches auf Erschütterungen und Schall reagieren kann. Die Geißel selbst ist unterschiedlich lang ausgebildet und kann auch sehr unterschiedlich geformt sein. Sie besitzt Sinneszellen, die vor allem der Geruchswahrnehmung dienen.
 Ernteameise im Rasterelektronenmikroskop
Ernteameise im RasterelektronenmikroskopAls drittes Segment folgt das Intercalarsegment, welches wiederum keine eigenen Strukturen trägt (es entspricht dem zweiten Antennensegment der Krebstiere). Am vierten bis sechsten Segment befinden sich die Mundwerkzeuge, beginnend mit den Mandibeln und den Maxillen, die jeweils paarig vorliegen, und abschließend mit dem Labium, das durch eine Verwachsung des Basalglieds unpaar ist und den Mundraum nach hinten abschließt. Den vorderen Abschluss bildet der Clypeus mit dem Labrum. Die Mandibel besteht nur aus einem Glied. Sie stellt als Beiß- und Kauwerkzeug bei den meisten Insekten das kräftigste Mundwerkzeug dar. Die Maxille lässt den Extremitätencharakter der Mundwerkzeuge sehr deutlich erkennen und erinnert an das Spaltbein der Krebstiere. Sie besteht aus einem Basalglied, dem Coxopoditen, der durch einen Cardo und einen Stipes geteilt ist. Darauf folgen zwei Kauladen, die als Galea und Lacinia bezeichnet werden, sowie ein mehrgliedriger Taster, der Palpus maxillaris. Das Labium entspricht diesem Aufbau, mit der Ausnahme, dass hier die beiden Coxopoditen verschmolzen sind und eine gemeinsame Basalplatte bilden, die in Mentum und Submentum unterteilt ist. Auch hier gibt es auf jeder Seite zwei Kauladen, die Glossae und die Paraglossae, sowie einen Palpus, den Palpus labialis. Vor allem die Palpen besitzen Sinneszellen zur Geruchs- und Geschmackswahrnehmung. Als weitere Strukturen des Mundraumes gibt es den Hypopharynx als zungenförmige Bildung des Mundvorraums und den Epipharynx an der Innenseite des Labrums.
Der hier beschriebene Bauplan entspricht dem Grundmuster der Insekten und wird als kauend-beißender Typ bezeichnet. Durch Abwandlungen der einzelnen Strukturen in Anpassung an unterschiedliche Ernährungsweisen können sich aus diesem Typus verschiedene andere Mundwerkzeugstypen entwickeln, darunter die leckend-saugenden Mundwerkzeuge vieler Fliegen oder die stechend-saugenden Mundwerkzeuge der Mücken oder der Wanzen.
Der Thorax
 Gewürfelte Tanzfliege (Empis tesselata)
Gewürfelte Tanzfliege (Empis tesselata) Behaarung einer Hummel
Behaarung einer HummelDer Thorax besteht aus drei Segmenten und trägt bei allen Insekten die sechs Laufbeine sowie bei den geflügelten Insekten zudem die beiden Flügelpaare. Die Einzelsegmente werden aufgrund ihrer Lage zueinander als Prothorax, Mesothorax und Metathorax bezeichnet. Jedes dieser Segmente trägt ein Paar der Thorakalbeine, die gemeinhin als Laufbeine ausgeprägt sind. Diese Beine können, abhängig von ihrer Funktion, sehr unterschiedlich gestaltet sein, besitzen jedoch immer den gleichen Grundaufbau. Sie bestehen aus einer Hüfte (Coxa), einem darauf folgenden Schenkelring (Trochanter), dem Oberschenkel (Femur), der Schiene (Tibia) sowie einem gegliederten Fuß (Tarsus), der aus einem bis fünf Tarsalgliedern sowie einem Praetarsus besteht. Der Praetarsus trägt im Normalfall zwei Krallen sowie bei vielen Insekten mehrere weitere Strukturen, die vor allem dem Festhalten an verschiedenen Oberflächen dienen.
Bei den Geflügelten Insekten tragen der Meso- und der Metathorax außerdem jeweils ein Paar Flügel, die bei den verschiedenen Insektengruppen unterschiedlich ausgeprägt sein können. Die ursprünglichen Insektengruppen wie die Felsenspringer und die Fischchen sind allerdings ungeflügelt, entsprechend sind Flügel nicht im Grundbauplan der Insekten vorhanden und erst später innerhalb der Insekten, mit der Entwicklung der Pterygota, entstanden. Stattdessen besitzen die Insekten im Grundbauplan seitlich an den Thoraxsegmenten Duplikaturen der Epidermis, die als Paranota bezeichnet und als Vorläuferstrukturen der Flügel angesehen werden.
Das Abdomen
Das Abdomen besteht ursprünglich aus elf Segmenten, von denen jedoch bei den verschiedenen Insektentaxa einzelne verschmolzen oder abhanden gekommen sind. Die Abdominalsegmente tragen nur abgewandelte Extremitäten und keine echten Beine. So finden sich etwa Styli, vor allem bei Larvenformen. Außerdem besitzt eine Reihe von Insektengruppen Cerci am letzten Segment, die für unterschiedliche Funktionen ausgebildet sind. Auch die Gonopoden, die die Begattungsorgane der Männchen und den Legeapparat (Ovipositor) der Weibchen bilden, sind umgewandelte Extremitäten. Das Abdomen enthält im Gegensatz zum Thorax nur sehr wenig Muskulatur, stattdessen sind hier die meisten Organsysteme lokalisiert.
Innere Anatomie und Physiologie
Nerven- und Hormonsystem
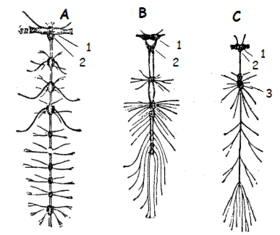 Nervensystem der Insekten (A – Termite, B – Schwimmkäfer, C – Fliege); 1 – Oberschlundganglion, 2 – Unterschlundganglion, 3 – anderer Nervenknoten
Nervensystem der Insekten (A – Termite, B – Schwimmkäfer, C – Fliege); 1 – Oberschlundganglion, 2 – Unterschlundganglion, 3 – anderer NervenknotenDas Nervensystem der Insekten entspricht im Wesentlichen dem gemeinsamen Grundbauplan der Mandibulata, zu denen neben ihnen auch die Krebstiere und Tausendfüßer gehören. Es besteht aus einem paarigen Bauchmarkstrang, der in jedem Segment ein Ganglienpaar aufweist. Im Kopf sind die Ganglien zu einem Oberschlundganglion, dem Gehirn, sowie zu einem Unterschlundganglion verschmolzen, beide sind miteinander über die Schlundkonnektive verbunden. Das Gehirn selbst weist einen sehr großen vorderen Bereich auf, das Protocerebrum, von dem beidseitig Bereiche in die Zentren der Komplexaugen ausstrahlen, die als Lobi optici bezeichnet werden. Auch die Ocellen werden von diesem Hirnteil innerviert. Vom Deuterocerebrum, dem zweiten Hirnabschnitt, ziehen Nerven in die Antennen und das Tritocerebrum versorgt den Vorderdarm durch eine Tritocerebralkommissur mit Nerven. Die Mundwerkzeuge werden von den im Unterschlundganglion konzentrierten Ganglien des vierten bis sechsten Segments versorgt. Durch den Thorax zieht sich das Bauchmark in Form eines typischen Strickleiternervensystems, wobei drei besonders große Ganglienpaare im Thorax die Beine und (wenn vorhanden) die Flügel versorgen. Das Abdomen enthält im Grundbauplan 7 normale Ganglienpaare und eine Ganglienmasse im achten Abdominalsegment, die aus den Ganglien aller folgenden Segmente besteht.
Das vegetative Nervensystem besteht aus drei Abschnitten. Dabei wird der vordere Abschnitt durch die Nerven des Mund- und Vorderdarmbereiches mit dem Frontal-, dem Hypocerebral- und dem Ventrikularganglion sowie den Corpora cardiaca und den Corpora allata gebildet. Den zweiten Abschnitt bildet die paarige Bauchganglienkette, der die Innervierung der Stigmen zukommt. Der hintere Darmabschnitt und die Genitalien werden vom caudalen Nerv versorgt. Besonders im Gehirn besitzen Insekten zudem neurosekretorische Drüsen, zu denen auch die bereits benannten Corpora cardiaca und Corpora allata gehören. Letztere schütten das Juvenilhormon aus, welches bei der Häutung das Entwicklungsstadium beeinflusst. Die Häutung selbst wird durch die Häutungshormone induziert, vor allem das Ecdyson.
Atmung
Als Atmungsorgane dienen bei den Insekten weitestgehend starre Röhren, die den gesamten Körper durchziehen und in immer feineren Röhren bis an die Organe und Einzelzellen reichen. Dieses System wird als Tracheensystem bezeichnet und entwickelt sich aus Einstülpungen der Epidermis nach innen. Entsprechend sind die Tracheen wie die Außenseite der Insekten mit einer Epidermis und einer dünnen Chitincuticula überzogen. Die Cuticula wird durch Taenidien versteift, spiralförmige Strukturen, die sich um die Röhre winden um ein Kollabieren zu verhindern. Nach außen öffnen sich die Tracheen in Atmungsöffnungen, die als Stigmen bezeichnet werden.
Der Atemvorgang ist primär passiv. Durch diese Form der Atmung ist auch bedingt, dass Insekten in wärmeren Gebieten größer werden können. In eher kalten Gebieten reicht die Molekularbewegung der Luft nicht aus, um die inneren Tracheen ausreichend mit Sauerstoff zu versorgen. Der Luftaustausch wird jedoch durch eine muskulöse Kontrolle der Stigmen reguliert und kann durch Ventilation erhöht werden. Bei vielen Insekten wurde die Anzahl der Stigmen von ursprünglich einem Paar an jedem Thorax- und Abdomensegment durch Querverbindungen (Anastomosen) reduziert, außerdem haben sich bei vielen Taxa Luftsäcke zur Unterstützung der Atmung und zur Sauerstoffspeicherung entwickelt. Bei Wasserinsekten haben sich zudem weitere Anpassungen entwickelt, die eine Atmung unter Wasser ermöglichen. So findet man bei vielen Wasserkäfern und anderen Wasserinsekten einen Bereich, der es durch eine besondere Oberfläche ermöglicht, eine Sauerstoffblase zu halten (Physikalische Kieme). Andere Arten besitzen ein volumenkonstantes Plastron oder Atemrohre zur Atmung. Vor allem bei wasserlebenden Insektenlarven wird auf die Tracheenatmung vollständig verzichtet und der Sauerstoff wird stattdessen über Tracheenkiemen oder die Haut aufgenommen.
Blutkreislauf
Das Blutgefäßsystem von Insekten besteht im Wesentlichen aus einem Hohlraum, dem Haemocoel, in dem alle inneren Organe in Blut (Hämolymphe) schwimmen (offener Blutkreislauf).
Da ein feines Blutgefäßsystem nicht benötigt wird, wurde dies auf ein einfaches Rückengefäß reduziert. Dieses besitzt einen kontraktilen Teil im Abdomen, der die Hämolymphe in den Körper pumpt und entsprechend Herz (nach caudal geschlossen) genannt wird, und einen Ast, der bis in den Kopf, das Gehirn und den Kopf mit Blut versorgt und Aorta (nach cranial offen) genannt wird. Im Kopf befindet sich ein sogenanntes „Antennenherz“, welches einen Muskel darstellt. Es „pumpt“ die Hämolymphe in die Antennenampullen und in die dazugehörigen Antennen. Der Blutstrom wird über die Peristaltik des Herzens in Bewegung gehalten, dabei wird die Hämolymphe von einem bis zwölf Paar seitlicher Öffnungen des Herzens, den Ostien, in das Herz „gesaugt“ und nach vorn durch die Aorta in den Kopf gepumpt. Von dort verteilt es sich über den Thorax in den Hinterleib und außerdem in die Beine und Flügel des Tieres. An den Beinen, Antennen und Flügeln sorgen kleinere Pumpsysteme für eine Versorgung bis in die Enden der Strukturen. Unterhalb des Herzens liegt ein dorsales Häutchen aus Muskulatur und Bindegewebe, welches als Diaphragma den Rückenteil des Körpers (Pericardialsinus) vom Hauptraum des Körpers (Perivisceralsinus) mit dem Darm und den Geschlechtsorganen trennt. Ein weiteres Diaphragma liegt oberhalb des Bauchteils (Perineuralsinus) mit dem Bauchmark.
Die Hämolymphe selbst macht 20 bis 40 Prozent des Körpergewichts aus und besteht aus Zellen, den Hämocyten, sowie dem flüssigen Plasma. Es transportiert das bei der Atmung entstehende Kohlendioxid, Proteine, Nährstoffe, Hormone und Exkrete und dient zudem der Osmoregulation und dem Aufrechterhalten des Körperinnendrucks. Das Blut ist meist farblos, aber es kann auch grünlich-gelb sein; es transportiert nicht nur Nährstoffe zu allen Körperteilen, sondern bringt auch die Hormone zu ihrem Bestimmungsort. Das Blut enthält spezielle Zellen, die Krankheitserreger bekämpfen, und andere, die Schäden oder Wunden reparieren. Anders als das Blut von Wirbeltieren enthält das Insektenblut keine Zellen, die mit der Atmung zu tun haben.
Die traditionelle Ansicht, das Blut der Insekten hätte aufgrund des effektiven Tracheensystems seine Funktion der Sauerstoffverteilung verloren, wurde in den letzten Jahren aufgrund neuer Befunde relativiert. Vorkommen des roten Blutfarbstoffs Hämoglobin sind bereits seit langem in aquatischen Insektenlarven, die unter Sauerstoffmangelbedingungen leben, wie z. B. Zuckmückenlarven, bekannt. Genomanalysen haben nun gezeigt, dass die Hämoglobinproduktion zum Grundvermögen beinahe aller Insekten gehört. Hämoglobin kommt vorwiegend in Kiemen und anderen respiratorischen Organen vor, so dass seine Funktion für die Atmung gesichert ist.[4] Bei zahlreichen Insektenordnungen wurde außerdem der blaue Blutfarbstoff Hämocyanin nachgewiesen, zumindest für eine Steinfliege wurde auch hier eine Funktion für die Sauerstoffversorgung nachgewiesen.[5]
Verdauung und Exkretion

Die Verdauung erfolgt im Darmsystem der Tiere. Dieses ist in drei funktionelle Abschnitte gegliedert, die entsprechend ihrer Lage als Vorder-, Mittel- und Enddarm bezeichnet werden. Der vordere und der hintere Anteil sind ektodermal gebildet und besitzen eine Auskleidung mit einer Chitincuticula, die entsprechend auch gehäutet werden muss. Der Vorderdarm beginnt mit der Mundöffnung und besteht vor allem aus dem muskulösen Pharynx. Über die Speiseröhre ist dieser Abschnitt mit dem Mitteldarm verbunden, wobei diese bei vielen Insekten auch Bereiche zur Nahrungsspeicherung (Ingluvies) oder einen Vormagen (Proventriculus) mit Kaustrukturen wie Leisten aus Chitin enthalten kann. Der Mitteldarm ist mit einem Drüsenepithel ausgekleidet und produziert die Enzyme, die zur Verdauung notwendig sind. Hier erfolgt auch die Resorption der Nährstoffe, die bei vielen Insekten in Blindschläuchen (Caeca) oder Krypten stattfindet. In den Caeca und Krypten können bei vielen Insekten auch symbiotische Mikroorganismen leben, die bei der Aufspaltung von bestimmten Nahrungsbestandteilen benötigt werden. Die unverdaulichen Reste (Exkremente) werden über den Enddarm ausgeschieden.
Die Exkretion der Insekten erfolgt über kleine Blindschläuche, die am Übergang des Mitteldarms zum Enddarm in den Darm münden. Diese werden als Malpighische Gefäße bezeichnet und sind wie der Enddarm ektodermalen Ursprungs. In den Zellen dieser Schläuche werden aktiv stickstoffhaltige Exkrete der Hämolymphe entzogen und mit den Exkrementen ausgeschieden. In den Rektalpapillen wird den Ausscheidungsprodukten vor der Ausscheidung noch Wasser entzogen.
Wichtige Strukturen der Nährstoff- und Exkretspeicherung sind die Fettkörper, die als große Lappen im Abdomen der Insekten liegen. Neben der Speicherung dienen sie der Synthese von Fetten und Glykogen sowie dem Abbau von Aminosäuren.
Geschlechtsorgane
Bis auf wenige Arten sind alle Insekten getrenntgeschlechtlich. Sehr wenige Arten sind Zwitter, eine Reihe von Arten pflanzt sich durch Parthenogenese fort.
 Paarung der Blauflügel-Prachtlibelle
Paarung der Blauflügel-PrachtlibelleDie Männchen besitzen paarige Hoden zur Spermienproduktion im Hinterleib, die über Samenleiter (Vasa deferentia) mit paarigen Samenbläschen (Vasa seminales) verbunden sind. Diese münden in einen unpaaren oder paarigen Ejakulationsgang, den Ductus ejaculatorius, und danach über mehr oder weniger komplex aufgebaute Begattungsorgane, den Aedoaegaus, meist im neunten Abdominalsegment nach außen. Zusätzlich können noch Zusatzdrüsen vorhanden sein, die Samenflüssigkeiten oder Stoffe zur Bildung von Spermatophoren bilden und den Spermien zugeben.
Auch die Ovarien der Weibchen sind im Regelfall paarig angelegt. Sie bestehen meistens aus einem Büschel einzelner Ovarienstränge, die als Ovariolen bezeichnet werden. Jede dieser Ovariolen besteht aus einem Germarium, in dem die Eizellen produziert werden, und einem Vittelarium zur Produktion der Dotterzellen. Abhängig von der Art, wie die Eier mit Dotter versorgt werden, unterscheidet man dabei drei verschiedene Formen von Ovariolen, die bei unterschiedlichen Insektentaxa vorkommen können. Bei der ersten Form, die als panoistische Ovariole bezeichnet wird, werden einzelne Eier im Vitellarium mit Dotter versorgt. Beim meroistisch-polytrophen Typ besitzt jede einzelne Eizelle mehrere Nährzellen und erhält über diese den Dotter. Bei der meroistisch-telotrophen Ovariole bleibt die einzelne Eizelle über einen Nährstrang mit dem Germarium verbunden und erhält den Dotter über diesen Weg. Um die heranwachsenden Eizellen legen sich bei allen Typen Follikelzellen.
Die Ovariolen vereinigen sich und enden in einer unpaaren Vagina, die zwischen dem siebten und neunten Abdominalsegment entweder direkt nach außen oder in eine Begattungstasche, der Bursa copulatrix, endet (Ausnahme: Eintagsfliegen). Im Bereich der Vagina ist bei fast allen Insekten eine Spermiensammeltasche, das Receptaculum seminis vorhanden, außerdem können verschiedene Anhangsdrüsen zur Produktion von Kittsubstanzen oder ähnlichem vorhanden sein.
Lebensräume

Insekten sind mit Ausnahme der Ozeane in fast allen Lebensräumen und Gebieten der Erde zu finden. Dabei existiert die größte Artenvielfalt in den tropischen Gebieten, während in Extremlebensräumen wie den Polargebieten, den Hochgebirgen und den küstennahen Meeresgebieten nur sehr wenige, aber hochangepasste Insektenarten leben. So findet man etwa in der Antarktis die Zuckmückenart Belgica antarctica, auf der Meeresoberfläche die zu den Wanzen zählenden Meerwasserläufer und im Gezeitenwatt die Larven von Zuckmücken der Gattung Clunio.
Einige Arten sind sehr stark spezialisiert und kommen entsprechend nur in besonders geeigneten Lebensräumen vor (stenöke Arten), andere dagegen können in fast allen Lebensräumen mit Ausnahme der Extremlebensräume leben (euryöke Arten) und wurden teilweise durch den Menschen weltweit verbreitet, so dass sie heute Kosmopoliten sind. Zu letzteren gehören vor allem verschiedene Arten der Schaben, Ameisen und Termiten sowie die als Nutztiere gehaltenen Honigbienen.
Die meisten Insekten leben in Böden oder auf bodennahen Strukturen sowie auf und in Pflanzen. Man geht davon aus, dass mit jeder Baumart der tropischen Regenwälder etwa 600 Insektenarten assoziiert sind, wobei man bei 50.000 Baumarten auf eine Artenzahl der Insekten von etwa 30 Millionen kommt. Auch auf Tierarten leben eine Reihe von Insekten, meistens als Ektoparasiten wie die verschiedenen Arten der Tierläuse und Flöhe oder als Kommensalen und Jäger im Fell der Tiere. Der Mensch stellt hierbei keine Ausnahme dar, auf ihm findet man etwa die verschiedenen Arten der Menschenläuse. Seltener sind Insekten Endoparasiten in Tieren. Zu nennen sind hier vor allem die zu den Zweiflüglern gehörenden Dasselfliegen, bei denen sich die Larven im Rachen (Rachendasseln), der Nasenhöhle (Nasendasseln) oder sogar im Magen (Magendasseln) von Pflanzenfressern entwickeln.
Lebensweise

Aufgrund ihrer Vielfalt haben Insekten heute beinah jede ihrer Größe angemessene ökologische Nische realisiert. Dabei spielt eine große Anzahl der Arten eine bedeutende Rolle bei der Remineralisierung organischer Stoffe im Boden, in der Bodenstreu, im Totholz und in anderen organischen Strukturen. Zu dieser Gruppe gehören auch die Leichenzersetzer, die in Tierleichen zu finden sind. Viele weitere Arten leben als Pflanzenfresser von lebenden Pflanzenteilen, das Spektrum reicht dabei von Wurzelhaaren über Holz bis hin zu Blättern und Blüten. Eine Reihe von Arten lebt als Nektar- und Pollensammler und spielt dabei eine wichtige Rolle bei der Pflanzenbestäubung. Wieder andere Insekten leben in und an Pilzen und ernähren sich von diesen. Eine große Gruppe von Insekten ernährt sich räuberisch von anderen Insekten oder kleineren Beutetieren. Eine letzte Gruppe stellen diejenigen Insekten dar, die sich von Teilen größerer Tiere wie Haaren, Schuppen und ähnlichem ernähren. In diese Gruppe gehören auch die zahlreichen Parasiten unter den Insekten, die beispielsweise Blut saugen oder sich in lebenden Geweben entwickeln.
Eine Besonderheit innerhalb der Insekten stellen verschiedene Arten von staatenbildenden Insekten dar. Diese Form des Zusammenlebens hat sich mehrfach unabhängig voneinander bei den Termiten und verschiedenen Hautflüglern (Ameisen, Bienen, Wespen) entwickelt. Bei diesen Tieren kommt es zum Aufbau eines Insektenstaates, in dem die Einzeltiere bestimmte Rollen innerhalb der Gesellschaft übernehmen. Häufig kommt es dabei zur Bildung von Kasten, deren Mitglieder sich morphologisch und in ihrem Verhalten gleichen. Bei vielen Ameisen findet man beispielsweise Arbeiter, Soldaten und Nestpfleger. Die Fortpflanzung übernehmen in diesen Fällen nur sehr wenige Geschlechtstiere innerhalb des Insektenstaates, manchmal nur eine einzige Königin, die befruchtete und unbefruchtete Eier legt.
Fortpflanzung und Entwicklung
 Wasserläufer bei der Paarung
Wasserläufer bei der PaarungDie Spermienübertragung erfolgt bei den Insekten ursprünglich über Spermatophoren, also Spermienpakete. Diese werden bei den Felsenspringern und den Fischchen von den Männchen auf dem Boden abgelegt und hier von den Weibchen aufgenommen. Bei allen folgenden Gruppen der Insekten gibt es eine direkte Spermienübertragung durch eine Kopulation, bei der die Spermien direkt in die Vagina oder die Bursa copulatrix eingebracht werden und hier entweder die Eizellen befruchten oder in das Receptaculum seminis zur Lagerung weitergeleitet werden. Die meisten Insekten legen nach der Begattung Eier ab (Ovoparie), andere sind ovovivipar, brüten die Eier also noch im Körper zur Schlupfreife aus. Noch seltener kommt es vor, dass bereits fertig entwickelte Larven (Larviparie) oder sogar Puppen (Pupiparie) zur Welt gebracht werden.
Die Furchung erfolgt bei den meisten Insekten superficiell. Das bedeutet, das sich auf dem sehr dotterreichen Ei mit zentralem Dotter (centrolecithales Ei) ein Furchungszentrum ausbildet, von dem die Furchung ausgeht. In seinem Bereich bilden sich mehrere Tochterkerne mit umgebenem Plasma (Furchungsenergiden), die durch Teilungen zu einem einschichtigen Blastoderm als Hüllepithel oder Serosa um den Dotter herum. Im ventralen Bereich bildet sich dann eine Keimanlage, die als Keimstreif in den Dotter hineinwächst und eine Höhle bildet (Amnionhöhle). In dieser Höhle findet die Hauptkeimbildung statt, nach deren Abschluss sich der Keim wieder nach außen entrollt und über dem Dotter der Rücken des Tieres geschlossen werden kann. Aus den Eiern schlüpfen dann Larven (Juvenilstadien mit eigenen Larvalmerkmalen) oder Nymphen (Juvenilstadien ohne eigene Merkmale).
 Hornisse, die aus ihrer Puppenhülle schlüpft
Hornisse, die aus ihrer Puppenhülle schlüpftAußerhalb des Eies folgt die postembryonale Entwicklung, nach der Insekten klassisch in hemimetabole und holometabole Insekten unterteilt werden. Dabei handelt es sich allerdings nur bei letzteren auch um eine taxonomische Gruppe, da die hemimetabole Entwicklung dem ursprünglichen Zustand entspricht. Die Entwicklung ist bei den verschiedenen Gruppen sehr unterschiedlich und hängt sehr stark von der Lebensweise der Juvenilstadien und der Imagines ab. Bei allen hemimetabolen Insekten kommt es über eine unterschiedliche Anzahl von Larven- oder Nymphenstadien ohne Puppenstadium zur Ausbildung des ausgewachsenen Tieres, der Imago. Zwischen den einzelnen Stadien findet dabei immer eine Häutung statt, bei der die alte Kutikula abgeworfen und eine neue angelegt wird. Abhängig von der Konzentration des Juvenilhormons im Blut kommt es dabei entweder zu einer Häutung von einer Larvenform in die nächste (bei viel Juvenilhormon) oder von einer Larvenform zur Imago (bei wenig Juvenilhormon). Den Zeitpunkt der Häutung bestimmt ein weiteres Hormon, das Ecdyson. Bei der Häutung kommt es vor allem zum Wachstum der Tiere, außerdem werden einzelne Merkmale neu angelegt. Dies geschieht durch eine Histolyse einzelner Strukturen und die Ausbildung von Imaginalanlagen oder das Ausstülpen spezieller Imaginalscheiben.
 Feuerwanzen, Larven und Imagines
Feuerwanzen, Larven und ImaginesBei den meisten hemimetabolen Insekten wie den verschiedenen Heuschreckenformen oder den Wanzen ähnelt die Nymphe in Grundzügen dem erwachsenen Tier und weist außer den fehlenden Flügeln keine besonderen Larvalanpassungen auf. Demgegenüber gibt es allerdings auch hemimetabole Insekten mit echten Larven, beispielsweise die Libellen oder die Eintagsfliegen. Eine feinere Unterteilung der Hemimetabolie ist möglich. So spricht man etwa von einer Palaeometabolie (Fischchen, Felsenspringer), wenn die Larven kaum eigene Merkmale aufweisen und sich nur durch Größenänderung zur Imago entwickeln. Dies ist auch bei den Eintagsfliegen der Fall, deren Imago zunächst keine Flügel besitzt und diese erst nach einer Imaginalhäutung erhält. Die Heterometabolie ist vor allem durch eine schrittweise Entwicklung der Flügel gekennzeichnet und kommt bei den Libellen, Steinfliegen und den meisten Schnabelkerfen vor. Schließlich gibt es noch die Neometabolie, bei der die Flügelanlagen erst bei den beiden letzten Larvenstadien angelegt werden; dies ist etwa bei einigen Zikaden und den Fransenflüglern der Fall.
Holometabole Insekten durchlaufen eine Metamorphose, ausgehend vom Ei über die Larve zur Puppe und dann zum erwachsenen Tier (Imago). Die Larve hat oft nicht die geringste physische Ähnlichkeit mit der Imago und besitzt eine Reihe von Eigenmerkmalen einschließlich anderer Lebensräume und Futterquellen im Vergleich zur Imago – ein nicht unerheblicher ökologischer Vorteil der Holometabola.
Populationsdynamik
Die Populationsdynamik vieler Insektenarten hat näherungsweise diskreten Charakter: In der zeitlichen Entwicklung einer Population können gegeneinander abgegrenzte, nicht-überlappende Generationen ausgemacht werden.
Schreibt man Nt für die Größe, oder nach Normierung Abundanz, einer Population zum Zeitpunkt t und r für die Anzahl der pro Individuum generierten Nachkommen, so ergibt sich für die einfachst mögliche Beschreibung einer Populationsdynamik:
Nt + 1 = rNt
Hängt der Anteil der selbst zur Reproduktion kommenden Nachkommen zum Zeitpunkt t+1 von der Größe der Population zum Zeitpunkt t ab, erhält man einen funktionalen Zusammenhang der Form:
Nt + 1 = f(Nt) mit einer Funktion f. Für f sind dabei verschiedene konkrete Ansätze möglich.Verwendung findet zum Beispiel die Hassel-Gleichung.
Auf einen ähnlichen Formalismus führt auch eine unmittelbare Abhängigkeit der Fertilität von der Populationsgröße.
Fossilbeleg

Das früheste einem Insekt sicher zuzuordnende Fossil ist Rhyniognatha hirsti aus dem Pragium, einer Stufe des Unterdevons vor etwa 407 Millionen Jahren. Aus derselben Formation stammt auch der älteste Springschwanz Rhyniella precursor, der als das älteste Insekt galt, solange diese zu den Insekten gerechnet wurden. Diese relativ fortgeschrittenen Fossilien lassen auf eine bereits länger dauernde Evolutionsgeschichte schließen, die vermutlich bis ins Silur zurückreicht. Insekten sind damit wohl nur unwesentlich jünger als die ältesten Funde von terrestrischen Tieren überhaupt. Da von Rhyniognatha bis auf die Kiefer nichts erhalten blieb, sind Aussagen darüber, wie dieses Insekt aussah und in welche heutige Verwandtschaft es einzuordnen wäre, spekulativ. Die Erstbeschreiber halten sogar einen Zusammenhang mit geflügelten Formen für denkbar, was allerdings sehr spekulativ bleibt.[6] Tatsächliche fossile Funde geflügelter Insekten liegen etwa von Gesteinen der Grenze Unterkarbon-Oberkarbon vor, das heißt, sie sind etwa 95 Millionen Jahre jünger.[7] (Ältere devonische Funde, als Eopterum devonicum beschrieben, erwiesen sich später als Reste des Carapax einer Krebsart.) Geflügelte Insekten liegen hier aus einer Vielzahl von Ordnungen, darunter sowohl lebende als auch ausgestorbene, vor. Die Flügelmorphologie erlaubte bereits unterschiedliche Flugstile.[8][9] Zur karbonischen Fauna gehörten z. B. die Palaeodictyoptera, die manchmal auch am ersten Thoraxsegment kleine flügelartige Fortsätze aufwiesen. Während des Karbons machten diese altertümlichen Ordnungen etwa ein Drittel der Insektenfauna aus; Schaben waren individuenreich vertreten, aber nicht sonderlich artenreich. Von den 27 im Karbon nachgewiesenen Insektenordnungen sind 8 am Übergang zum Perm oder wenig später ausgestorben. Drei weitere Ordnungen starben am Übergang zur Trias aus. Seit diesem evolutionären „Flaschenhals“ haben alle überlebenden Linien bis heute mehr oder weniger stetig an Diversität zugenommen, auch das dritte Massenaussterben an der Kreide/Tertiär-Grenze scheint die Insekten nur wenig betroffen zu haben.[10] Wichtiger für diese erhöhte Diversität scheinen überraschenderweise nicht schnellere Artbildungsraten, sondern verminderte Aussterberaten zu sein.[11]
Sowohl Radiationen wie längere Perioden mit niedrigen Artenzahlen und wenig evolutionären Neuerungen können mit ausgeprägten Schwankungen des Sauerstoffspiegels der Erde im Paläozoikum in Verbindung gebracht werden, ausgedehnte Perioden mit sehr geringen fossilen Funden sind daher wohl nicht ausschließlich auf Zufälle der fossilen Überlieferung zurückzuführen, sondern können durchaus real sein.[12] Während der Perioden mit ungewöhnlich hohem Sauerstoffgehalt lebten spektakuläre Rieseninsekten mit Körpergrößen, die alle rezenten Formen bis um das Zehnfache übersteigen.[13] Die Urlibelle Meganeura monyi (gefunden in Commentry, Frankreich) erreichte eine Flügelspannweite von 63 cm,[14] Meganeuropsis permiana soll sogar 71 cm Spannweite besessen haben, wahrscheinlich nahe an der Obergrenze des Arthropoden-Bauplans für Flugfähigkeit.[15]
Im Perm tauchten die meisten modernen Insektenordnungen auf, doch spielten gerade die heute dominierenden holometabolen Insekten lange Zeit eine untergeordnete Rolle,[16] und erst ab der Trias kam es bei diesen zu einer wahren Explosion der Arten- und Formenvielfalt. Gründe für diesen Erfolg werden im Massenaussterben am Ende des Perms gesehen (bisher dominierende Insekten starben aus), in besserer Widerstandsfähigkeit gegen das aride Klima des Perms und in der Ausbreitung der Samenpflanzen.[17]
Alle gegenwärtig anerkannten Insektenordnungen und knapp zwei Drittel der Familien sind auch fossil belegt, wobei ihre tertiäre Artenzahlen gut mit ihren heutigen korrelieren (Ausnahme: Schmetterlinge, die fossil weniger artenreich belegt sind).
Systematik
Drei Gruppen, die traditionell als Urinsekten zu den Insekten gezählt wurden, die Springschwänze (Collembola), Doppelschwänze (Diplura) und Beintastler (Protura) gelten heute nicht mehr als eigentliche Insekten. Sie werden zusammen mit diesen hier innerhalb der übergeordneten Gruppe der Sechsfüßer (Hexapoda) geführt. Sowohl die Hexapoda als auch die Insekten als solche gelten aufgrund ihrer typischen Merkmale (Apomorphien) als gesicherte Taxa. Die tatsächliche Schwestergruppe der Insekten innerhalb der Hexapoda ist allerdings umstritten.
Klassische Systematik
Die Insekten werden häufig auch als Freikiefler (Ectognatha) den Sackkieflern (Entognatha) gegenüber gestellt, die aus eben den benannten Taxa der Urinsekten bestehen. Sie besitzen als wichtigste Eigenmerkmale eine Geißelantenne mit einem Grundglied (Scapus) und einem Pedicellus mit Johnstonschem Organ. Die eigentliche Geißel ging auf das ursprünglich dritte Glied der Antenne zurück, welches sich in eine unterschiedliche Anzahl von Geißelsegmenten aufgeteilt hat. Weitere Apomorphien sind der Besitz rückwärtiger Äste des Tentoriums, einer Skelettstruktur im Kopf, paarige Krallen am Praetarsus mit einer gelenkigen Verbindung zu selbigem. Auch der primär in fünf Tarsenglieder gegliederte Tarsus und die Umbildung der hinteren Abdominalextremitäten zu Gonopoden und die Beteiligung derselben am Legeapparat (Ovipositor) der Weibchen wird als neu erworbenes Merkmal angesehen. Das ursprünglich namensgebende Merkmal, die freiliegenden Kiefer, ist dagegen ein ursprüngliches Merkmal (Plesiomorphie), die bereits bei den gemeinsamen Vorfahren der Krebstiere, Hundertfüßer, Tausendfüßer und Hexapoda vorhanden war (Mandibulata).
Innerhalb der Insekten werden in der klassischen Systematik heute je nach Quelle unterschiedliche Anzahlen von Unterklassen, Überordnungen und Ordnungen unterschieden. Dies hängt mit der in Teilen noch unverstandenen tatsächlichen Verwandtschaft der Insektengruppen untereinander sowie der unterschiedlichen Ansicht der Beschreiber über die Anordnung innerhalb einer klassischen Hierarchiebeschreibung zusammen. In diesem Werk soll folgender Systematik gefolgt werden, Unterschiede zu anderen Werken ergeben sich vor allem im Rang der Libellen und Eintagsfliegen, die häufig als Ordnung eingeordnet werden, sowie bei den Schnabelkerfen, deren Unterordnungen in einigen Werken als eigenständige Ordnungen geführt werden, ebenso bei den Heuschrecken, bei denen häufig noch die Kurzfühlerschrecken und die Langfühlerschrecken als Unterordnungen der Ordnung Orthoptera (syn. Saltatoria) eingestuft werden, wie z. B. in Fauna Europaea oder Orthoptera Species File Online.
- Unterklasse Felsenspringer (Archaeognatha)
- Unterklasse Fischchen (Zygentoma)
- Unterklasse Fluginsekten (Pterygota)
- Überordnung Eintagsfliegen (Ephemeroptera)
- Überordnung Libellen (Odonata)
- Überordnung Neuflügler (Neoptera)
- Ordnung Steinfliegen (Plecoptera)
- Ordnung Tarsenspinner (Embioptera)
- Ordnung Grillenschaben (Notoptera)
- Ordnung Ohrwürmer (Dermaptera)
- Ordnung Fangschrecken (Mantodea)
- Ordnung Schaben (Blattodea)
- Ordnung Termiten (Isoptera)
- Ordnung Gespenstschrecken (Phasmatodea)
- Ordnung Gladiatoren (Mantophasmatodea)
- Ordnung Langfühlerschrecken (Ensifera)
- Ordnung Kurzfühlerschrecken (Caelifera)
- Ordnung Bodenläuse (Zoraptera)
- Ordnung Staubläuse (Psocoptera)
- Ordnung Tierläuse (Phthiraptera)
- Ordnung Fransenflügler (Thysanoptera)
- Ordnung Schnabelkerfe (Hemiptera)
- Unterordnung Rundkopfzikaden (Cicadomorpha)
- Unterordnung Spitzkopfzikaden (Fulgoromorpha)
- Unterordnung Wanzen (Heteroptera)
- Unterordnung Scheidenschnäbler (Coleorrhyncha)
- Unterordnung Pflanzenläuse (Sternorrhyncha)
- Ordnung Kamelhalsfliegen (Raphidioptera)
- Ordnung Großflügler (Megaloptera)
- Ordnung Netzflügler (Neuroptera)
- Ordnung Käfer (Coleoptera)
- Ordnung Fächerflügler (Strepsiptera)
- Ordnung Hautflügler (Hymenoptera)
- Ordnung Köcherfliegen (Trichoptera)
- Ordnung Schmetterlinge (Lepidoptera)
- Ordnung Schnabelfliegen (Mecoptera)
- Ordnung Flöhe (Siphonaptera)
- Ordnung Zweiflügler (Diptera)
Phylogenetische Systematik
Die phylogenetische Systematik versucht, das System anhand der tatsächlichen Verwandtschaft aufzubauen. Unterschiede zur klassischen Systematik ergeben sich vor allem in der Behandlung der sog. paraphyletischen Gruppen. Paraphyletische Taxa werden nur durch altertümliche Merkmale Symplesiomorphien abgegrenzt. Sie umfassen nicht alle Nachfahren eines tatsächlich gemeinsamen Vorfahren. Wichtigste Methode ist nach wie vor eine sorgfältige Untersuchung der Anatomie und Morphologie, auch unter Einschluss aussagekräftiger Fossilien.
Als zusätzliche Methode sind heute molekulare Stammbäume von sehr großer Bedeutung. Molekulare Stammbäume beruhen auf dem direkten Vergleich homologer DNA-Sequenzen. Dazu werden dieselben Genabschnitte der verschiedenen zum Vergleich stehenden Organismen mit Hilfe der Polymerase-Kettenreaktion vervielfältigt, sequenziert und anschließend verglichen. Mittels verschiedener statistischer Techniken werden die Sequenzen zu einem molekularen Stammbaum angeordnet, der im Idealfall genau die abgestufte Verwandtschaft der untersuchten Individuen widerspiegelt. Häufig verwendete Gene für den molekularen Stammbaum der Insekten sind Gene für Ribosomale RNA, Histon H3, Elongationsfaktor 1 alpha 1 und verschiedene mitochondriale Gene (z. B. mt-CO1), zunehmend aber auch die gesamte Mitochondriale DNA. Ein Vergleich auf Basis des gesamten sequenzierten Genoms wäre naturgemäß am aussagekräftigsten, er wurde aufgrund der wenigen vollständig sequenzierten Insektengenome auch tatsächlich bereits unternommen.[18] Wie jede andere Methode hat auch die Erstellung molekularer Stammbäume ihre Schwierigkeiten und Fallstricke. Wie häufig bei neuen Methoden, ist die anfängliche Euphorie einer gewissen Ernüchterung gewichen, weil zu viele an verschiedenen Genen oder mit verschiedenen Methoden erhobene Stammbäume einander krass widersprechende Ergebnisse erbrachten.[19][20] Ein Hauptproblem besteht darin, dass viele Insektenordnungen offensichtlich in einer schnell ablaufenden Radiation in kurzer zeitliche Folge entstanden sind und sich anschließend über mehr als dreihundert Millionen Jahre auseinanderentwickelten, wodurch die Informationen der ursprünglichen Radiation kaum noch aufzuschlüsseln sein werden.[21][22]
Aktuelle Vorschläge für phylogenetische Systematiken beruhen in der Regel auf einer Kombination von morphologischen und molekularen Stammbäumen. Zusätzlich werden sog. "supertrees" aufgestellt, die die Ergebnisse zahlreicher Studien zusammenführen, ohne selbst neue Daten beizutragen. Auch die zur Zeit intensiv diskutierten Stammbäume (u. a.:[23][24][25][26]) kommen an zahlreichen Punkten nicht zur Übereinstimmung. Dennoch haben sich gegenüber früheren auf rein morphologischer Basis erstellten Systematiken (klassische Grundlage:[27]) zahlreiche Verbesserungen ergeben.
Das phylogenetische System kommt ohne die Rangstufen der klassischen Systematik oberhalb der Art aus. In der Regel werden die Insektenordnungen aber als Verwandtschaftsgruppen beibehalten, auch wenn klar ist, dass sie weder gleiches Alter noch gleichen tatsächlichen Rang besitzen können. Folgende klassische Ordnungen haben sich unzweifelhaft als paraphyletisch erwiesen:[28]
- Schaben, in Bezug auf Termiten (wahrscheinlichste Schwestergruppe: Gattung Cryptocercus)
- Staubläuse, in Bezug auf Tierläuse (wahrscheinlichste Schwestergruppe: Bücherläuse (Liposcelidae)).
- Schnabelfliegen, in Bezug auf Flöhe (wahrscheinlichste Schwestergruppe: Winterhafte (Boreidae)).
Die jeweils zweitgenannte Gruppe ist also in Wirklichkeit keine Schwestergruppe der erstgenannten, sondern ein Teil davon (d. h., sie ist mit einigen Familien davon näher verwandt, als diese es untereinander sind). Die Monophylie der übrigen Ordnungen wurde, manchmal nach zwischenzeitlich aufgekommenen Zweifeln, bestätigt (v. a. Schnabelfliegen in Bezug auf Zweiflügler und Großflügler in Bezug auf Kamelhalsfliegen: noch nicht sicher).
Das im folgenden dargestellte System (nach den o. g. Quellen kombiniert) ist unsicher, an verschiedenen Stellen umstritten und wird sich möglicherweise in den nächsten Jahren noch verändern, es gibt in etwa den heutigen Stand des Wissens wieder. Die Verwandtschaftsverhältnisse innerhalb der Polyneoptera (deren Monophylie heute fast unstrittig ist) weiter aufzulösen, ist zur Zeit mit hinreichender Sicherheit unmöglich. Einige traditionell sehr umstrittene Fragen, wie die Stellung der Fächerflügler oder der Bodenläuse, scheinen aber einer Lösung nähergerückt.
Insekten Dicondylia Fluginsekten Metapterygota Neuflügler Polyneoptera Dictyoptera Notoptera N.N. Springschrecken Eumetabola Paraneoptera Staubläuse und Tierläuse
Holometabola N.N. Neuroperiforma Neuropterida Neuroperiforma Coleopteroida Mecopteroida Amphiesmenoptera Antliophora N.N. Insekten und Menschen
Insekten als Schädlinge
 Anopheles bei der Blutmahlzeit
Anopheles bei der BlutmahlzeitEine Reihe von Insekten ist als Schädling (Ungeziefer) an Nutz- und Zierpflanzen, Holzkonstruktionen und -produkten (Holzschutz) und Nahrungsvorräten bekannt, andere leben als Parasiten am Menschen und seinen Haustieren. Ferner sind einige Insektenarten, vor allem in großen Individuenzahlen vorkommende Blutsaugende Insekten, Überträger (Vektoren) von Krankheiten wie der Rattenfloh für die Pest, Mücken der Gattung Anopheles für die Malaria oder die Tse-Tse-Fliegen für die Schlafkrankheit.
Der Befall mit Pflanzenschädlingen kann bei der heute üblichen Monokultur von Nutzpflanzen zu großen Ausfällen der Ernte führen. Neben der direkten Schädigung durch Fraß übertragen viele Insekten auch Pflanzenkrankheiten, meist Virosen und Pilzerkrankungen. Viele weitere Schädlinge leben als Vorratsschädlinge auch in geernteten oder auch bereits verarbeiteten Materialien und führen hier zu weiteren Schäden. Dem stehen andere Insektenarten gegenüber, die im Sinne der biologischen Schädlingsbekämpfung eingesetzt werden und Schädlinge direkt jagen oder in ihnen ihre Eier ablegen. Vor allem verschiedene Schlupfwespenarten werden entsprechend gezielt gegen spezifische Schädlinge eingesetzt.
Nutzung von Insekten
Als Nutztiere hält der Mensch vor allem die Honigbiene, die Schlupfwespen zur biologischen Schädlingsbekämpfung, die Raupen des Seidenspinners sowie in vielen Ländern Heuschrecken und Grillen als Nahrungsmittel. Der Verzehr von Insekten ist über große Teile Afrikas, Südostasiens und Mittel- bzw. Südamerikas verbreitet und wird als Entomophagie bezeichnet. Dabei stellen Insekten eine proteinreiche Kost dar; etwa 500 Arten werden weltweit für die menschliche Ernährung genutzt. Allerdings spielen auch sehr viele Insekten sekundär als Pflanzenbestäuber eine große Rolle in der Nahrungsproduktion des Menschen, da ohne sie keine Fruchtbildung stattfinden könnte.
Einige Insekten werden auch als Haustiere in Terrarien gehalten. Besonders zu nennen sind dabei die Ameisen, die in Formicarien gehalten werden. Sie gehören zu den Hautflüglern und sind somit enge Verwandte der Bienen, Wespen und Hornissen. Weitere beliebte Insekten der Terrarianer sind Stabheuschrecken, Fangschrecken und verschiedene Käfer- und Schabenarten, außerdem werden Mehlkäferlarven (Mehlwürmer), Grillen und andere Insekten als Futtertiere für Reptilien und als Angelköder gezüchtet. Eine Entwicklung der letzten Jahrzehnte ist die Haltung von Schmetterlingen in großen Gewächshäusern, die als Schmetterlingshäuser für Besucher geöffnet werden und an manchen Orten zu den Sehenswürdigkeiten der Region werden.
Auch in der pharmazeutischen Industrie werden verschiedene Insekten eingesetzt, das bekannteste Beispiel stellen hierbei die Spanische Fliege (Lytta vesicatoria) sowie einige weitere Ölkäfer dar, die den Stoff Cantharidin produzieren. Als wissenschaftliche Versuchstiere haben sich Drosophila melanogaster sowie verschiedene Heuschrecken- und Käferarten etabliert. Die Leichenzersetzer unter den Insekten, vor allem die Larven verschiedener Fliegen und Käfer, spielen heute zudem eine bedeutende Rolle in der Kriminalistik. Der Forschungszweig der Entomologischen Forensik zur Aufklärung von Kriminalfällen basiert auf der Erforschung dieser Tiere. Schmeißfliegenlarven werden in der Wundheilung eingesetzt. Außerdem werden verschiedene Arten der Schildläuse zur Produktion von Farbstoffen, Lacken oder Wachsen genutzt wie z. B. Karmin (Cochenille) und Schellack.
Als sogenannte Insekten-Cyborgs sollen mit militärischer Technik schon während der Metamorphose ausgerüstete Insekten Dienst für die amerikanischen Streitkräfte leisten: „DARPA seeks innovative proposals to develop technology to create insect-cyborgs, possibly enabled by intimately integrating microsystems within insects, during their early stages of metamorphoses.“( [29])
Materialien zu Insekten
Sich reinigende Fleischfliege (4:05 Minuten Film) – 6 MB Xvid in Ogg-Container
Der Film zeigt eine Fleischfliege in Portugal, die mit Hilfe ihrer Vorder- und Hinterbeine ihre Flügel und ihren Kopf reinigt. Um die sehr schnellen Bewegungen der Fliege besser sehen zu können, wird der Film mit halber Geschwindigkeit wiedergegeben.Museen/Sammlungen
Zur Erfassung der Artenvielfalt und der Variationsbreite der Insekten werden Insektensammlungen von Privatsammlern und wissenschaftlichen Einrichtungen angelegt. Vor allem Museensammlungen erreichen dabei beachtliche Größen und Individuenzahlen. Die wissenschaftliche Insektensammlung im Naturhistorischen Museum des Stiftes Admont (Steiermark) von Pater Gabriel Strobl (1846–1925) beinhaltet beispielsweise etwa 252.000 Exemplare aus 57.000 verschiedenen Arten, die Dipteren-Sammlung zählt mit ihren etwa 80.000 aufbewahrten Exemplaren und rund 7500 verschiedenen Artnamen zu den drei bedeutendsten Fliegen-Kollektionen in Europa.
Auch private Sammler legen sich häufig Insektensammlungen an; vor allem auffällige Schmetterlinge und Käfer werden dabei gerne aufgrund ästhetischer Vorlieben oder seltene Arten aufgrund ihres Seltenheitswertes gesammelt. Von einer Sammlung aus rein ästhetischen Gründen ist abzuraten, da vor allem die Populationen seltener Schmetterlinge wie des europäischen Apollofalters durch die Sammelleidenschaft (neben ihrem Lebensraumverlust und anderer Gründe) an den Rand der Ausrottung gebracht werden können. Auf der anderen Seite gibt es vor allem in tropischen Ländern wirtschaftliche Interessen an der vor allem in Europa und Nordamerika beliebten Sammelleidenschaft. Dort werden Schmetterlinge und andere beliebte Insekten in speziellen Farmen gezüchtet und an Sammler verschickt.
Kulturelle Bedeutung
In die Kultur des Menschen haben vor allem die ihm nützlichen Insektenarten wie die Honigbiene und andere in seinem direkten Umfeld lebende Arten Einzug gefunden. Die bekannteste Verarbeitung des Insektenmotivs in der Literatur ist dabei Die Biene Maja von Waldemar Bonsels. In der Musik sind das mexikanische Volkslied La Cucaracha und Der Hummelflug besonders populär. Letzterer fand über die Band Manowar auch Einzug in den Heavy Metal. In Deutschland ist das Kinderlied „Maikäfer flieg!“ lange Zeit kulturelles Allgemeingut gewesen.
Siehe auch
Literatur
- Michael Chinery: Field Guide to Insects of Britain and Northern Europe. 3. Auflage. Harper Collins, 1993, 448 S., ISBN 0-00219-918-1
- Michael Chinery: Pareys Buch der Insekten. Franckh-Kosmos, o. O. 2004, 328 S., ISBN 3440099695
- Bernhard Klausnitzer: Insecta (Hexapoda), Insekten, in: W. Westheide, R. Rieger (Hrsg.): „Spezielle Zoologie Teil 1: Einzeller und Wirbellose Tiere“; Gustav Fischer Verlag, Stuttgart, Jena.
- Christopher O’Toole: Faszinierende Insekten. Bechtermünz Verlag, 223 S., ISBN 3-8289-1584-1
- E. Stresemann (Begr.), H.-J. Hannemann, B. Klausnitzer, K. Senglaub: Exkursionsfauna von Deutschland, Wirbellose: Insekten. 9. Aufl. Spektrum Akademischer Verlag, Heidelberg, Berlin 2000, 959 S., ISBN 3-8274-0922-5
Einzelnachweise
- ↑ David Grimaldi & Michael S. Engel (2005): Evolution of the Insects. Cambridge University Press ((Cambridge Evolution Series) ISBN 0521821495.
- ↑ Ebbe Schmidt Nielsen & Laurence A. Mound (1997): Global Diversity of Insects: The Problems of Estimating Numbers. In: Peter H. Raven & Tania Williams (Hrsg.): Nature and Human Society. The Quest for a Sustainable World. Proceedings of the 1997 Forum on Biodiversity. (National Research Council USA, Board on Biology)
- ↑ neuere Literatur zusammengefasst in: Gerhard Scholtz & Gregory D. Edgecombe (2006): The evolution of arthropod heads: reconciling morphological, developmental and palaeontological evidence. Development Genes and Evolution Volume 216, Numbers 7-8: 395-415, doi:10.1007/s00427-006-0085-4
- ↑ Thorsten Burmester & Thomas Hankeln (2007): The respiratory proteins of insects. Journal of Insect Physiology Volume 53, Issue 4: 285-294. doi:10.1016/j.jinsphys.2006.12.006
- ↑ Christian Pick, Marco Schneuer, Thorsten Burmester (2009): The occurrence of hemocyanin in Hexapoda. FEBS Journal 276: 1930–1941. doi:10.1111/j.1742-4658.2009.06918.x
- ↑ Michael S. Engel & David A. Grimaldi (2004): New light shed on the oldest insect. Nature 427, 627-630. doi:10.1038/nature02291
- ↑ Carsten Brauckmann, Brigitte Brauckmann, Elke Gröning (1994): The stratigraphical position of the oldest known Pterygota (insecta. Carboniferous, Namurian). Annales de la Société Géologique de Belgique 117 (Hommage à Maurice Streel) Fascicule 1: 47-56.
- ↑ Robin J. Wootton (2003): Reconstructing insect flight performance from fossil evidence. Acta zoologica cracoviensia 46 (suppl.– Fossil Insects): 89-99.
- ↑ R. J. Wootton & J. Kulakova-Peck (2000): Flight adaptations in Palaeozoic Palaeoptera (Insecta). Biological Reviews 75: 129–167. doi: 10.1111/j.1469-185X.1999.tb00043.x
- ↑ Paul Whalley (1987): Insects and Cretaceous mass extinction. Nature 327: 562 doi:10.1038/327562b0
- ↑ Conrad C. Labandeira & J. John Seposki Jr.(1993): Insect diversity in the fossil record. Science 261 (5119): 310-315.
- ↑ Peter Ward, Conrad Labandeira, Michel Laurin, Robert A. Berner (2006): Confirmation of Romer's Gap as a low oxygen interval constraining the timing of initial arthropod and vertebrate terrestrialization. PNAS vol. 103 no. 45: 16818-16822. doi:10.1073/pnas.0607824103
- ↑ Jon F. Harrison, Alexander Kaiser, John M. VandenBrooks (2010): Atmospheric oxygen level and the evolution of insect body size. Proceedings of the Royal Society Series B (2010) 277: 1937–1946. doi:10.1098/rspb.2010.0001
- ↑ F. M. Carpenter (1960): Studies on North American Carboniferous insects. 1. The Protodonata. Psyche, 67: 98- 110.
- ↑ William A. Shear & Jarmila Kulakova-Peck (1990): The ecology of Paleozoic terrestrial arthropods: the fossil evidence. Canadian Jounal of Zoology 68: 1807- 1834.
- ↑ nach neueren Funden existieren holometabole Insekten seit dem Oberkarbon, waren anfangs aber offensichtlich sehr selten: André Nel, Patrick Roques, Patricia Nel, Jakub Prokop, J. Sébastien Steyer (2007): The earliest holometabolous insect from the Carboniferous: a “crucial” innovation with delayed success (Insecta Protomeropina Protomeropidae). Annales de la Société entomologique de France (n.s.) 43(3): 349-355
- ↑ Olivier Béthoux: The earliest beetle identified. In: Journal of Paleontology. 6, Nr. 83, 2009, S. 931–937, doi:10.1666/08-158.1.
- ↑ Evgeny M. Zdobnov & Peer Bork (2007): Quantification of insect genome divergence. Trends in Genetics Vol.23 No.1 : 16-20. doi:10.1016/j.tig.2006.10.004
- ↑ Cameron, S. L., Miller, K. B., D'Haese, C. A., Whiting, M. F. and Barker, S. C. (2004): Mitochondrial genome data alone are not enough to unambiguously resolve the relationships of Entognatha, Insecta and Crustacea sensu lato (Arthropoda). Cladistics, 20: 534–557. doi:10.1111/j.1096-0031.2004.00040.x
- ↑ Bitsch, J., Bitsch, C., Bourgoin, T., D'Haese, C. (2004): The phylogenetic position of early hexapod lineages: morphological data contradict molecular data. Systematic Entomology 29: 433–440. doi:10.1111/j.0307-6970.2004.00261.x
- ↑ James B. Whitfield & Karl M. Kjer (2008): Ancient Rapid Radiations of Insects: Challenges for Phylogenetic Analysis. Annual Revue of Entomology 53: 449–472. doi:10.1146/annurev.ento.53.103106.093304
- ↑ K.D.Klass (2007): Die Stammesgeschichte der Hexapoden: eine kritische Diskussion neuerer Daten und Hypothesen. Denisia 20: 413-450. PDF
- ↑ Beutel, R. G., Friedrich, F., Hörnschemeyer, T., Pohl, H., Hünefeld, F., Beckmann, F., Meier, R., Misof, B., Whiting, M. F., Vilhelmsen, L. (2011): Morphological and molecular evidence converge upon a robust phylogeny of the megadiverse Holometabola. Cladistics, 27: 341–355. doi:10.1111/j.1096-0031.2010.00338.x
- ↑ Jerome C. Regier, Jeffrey W. Shultz, Andreas Zwick, April Hussey, Bernard Ball, Regina Wetzer, Joel W. Martin, Clifford W. Cunningham (2010): Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences. Nature 463: 1079-1083 doi:10.1038/nature08742
- ↑ Keisuke Ishiwata, Go Sasaki, Jiro Ogawa, Takashi Miyata, Zhi-Hui Sua (2011): Phylogenetic relationships among insect orders based on three nuclear protein-coding gene sequences. Molecular Phylogenetics and Evolution Volume 58, Issue 2: 169-180 doi:10.1016/j.ympev.2010.11.001
- ↑ Robert B. Davis, Sandra L. Baldauf, Peter J. Mayhew (2010): Many hexapod groups originated earlier and withstood extinction events better than previously realized: inferences from supertrees. Proceedings of the Royal Society London Series B online (Downloaded from rspb.royalsocietypublishing.org on August 18, 2011) doi:10.1098/rspb.2009.2299
- ↑ Willi Hennig (1969): Die Stammesgeschichte der Insekten. Kramer, 1969.
- ↑ umfassende Darstellung z. B. in: Jan Zrzavy (2008): Four chapters about the monophyly of insect ‘orders’: A review of recent phylogenetic contributions. Acta entomologica musei nationalis Pragae Volume 48(2): 217-232. download vom Herausgeber unter PDF
- ↑ BAA 06-22 - Solicitations - Microsystems Technology Office
Weblinks
 Commons: Insekten – Sammlung von Bildern, Videos und Audiodateien Commons: Insekten als Thema – Sammlung von Bildern, Videos und Audiodateien
Commons: Insekten – Sammlung von Bildern, Videos und Audiodateien Commons: Insekten als Thema – Sammlung von Bildern, Videos und Audiodateien Wiktionary: Insekt – Bedeutungserklärungen, Wortherkunft, Synonyme, Übersetzungen
Wiktionary: Insekt – Bedeutungserklärungen, Wortherkunft, Synonyme, Übersetzungen- ausführliche Insektenseite
- Systematik, Körperbau, Fotos und Angaben zur Lebensweise von über 850 Insektenarten in Mitteleuropa
- Bugbios – Insects on the Web, englisch
- Insektenbestimmung
- Fotos fliegender Insekten, aufgenommen in freier Natur
- Insect Evolution Kommentiertes Link-Verzeichnis (in Englisch)


Dieser Artikel wurde am 6. Januar 2006 in dieser Version in die Liste der lesenswerten Artikel aufgenommen. Kategorien:- Wikipedia:Lesenswert
- Insekten


















Wikimedia Foundation.