- Krallenträger
-
Stummelfüßer 
Stummelfüßer in der Region Bay of Plenty im Norden der neuseeländischen Nordinsel
Systematik Unterreich: Vielzellige Tiere (Metazoa) Abteilung: Gewebetiere (Eumetazoa) Unterabteilung: Bilateria ohne Rang: Urmünder (Protostomia) Überstamm: Häutungstiere (Ecdysozoa) Stamm: Stummelfüßer Wissenschaftlicher Name Onychophora Familien Die Stummelfüßer (Onychophora - wörtlich Krallenträger) bilden einen eigenen Tierstamm aus der Gruppe der Häutungstiere (Ecdysozoa) und lassen sich vereinfacht als Würmer mit Beinen beschreiben. Hauptsächlich auf der Südhalbkugel verbreitet, leben sie räuberisch von kleineren Tieren wie zum Beispiel Insekten, die sie oft durch Verspritzen eines klebrigen Schleims erbeuten. In der modernen Zoologie sind sie nicht zuletzt durch die lebendgebärende Fortpflanzungsweise und die ausgefallenen Begattungsformen vieler Arten bekannt. Mögliche Vorfahren der Tiere, die Lobopoden aus der erdgeschichtlichen Epoche des Kambriums, sind in der Paläontologie von großem Interesse.
Stummelfüßer sind sehr wahrscheinlich nahe mit Gliederfüßern (Arthropoda) und Bärtierchen (Tardigrada) verwandt und bilden mit diesen beiden das Taxon Panarthropoda. Die erste Gattung wurde 1825 von Landsdown Guilding wissenschaftlich beschrieben, der die Tiere noch als abgewandelte Schnecken (Gastropoda) ansah; der Name Onychophora wurde erst 1853 geprägt.
Inhaltsverzeichnis
Aufbau
Stummelfüßer sind wurmförmige, segmentierte Tiere mit zylindrischem, bauchseitig abgeflachten Körperquerschnitt und reihenförmig angeordneten ungegliederten Körperanhängen, den Stummelfüßen. Die Tiere werden zwischen 0,5 und 20 Zentimeter lang, wobei der Durchschnitt bei etwa 5 Zentimetern liegt, und weisen zwischen 13 und 43 Beinpaare auf. Die Farbe ihrer von zahlreichen feinen Querringen bedeckten Haut ist oft unauffällig orangefarben, rot oder braun, manchmal aber auch leuchtend grün, blauschwarz, goldfarben oder weiß, gelegentlich auch farbig gemustert.
Die äußerlich unauffällige und nur am regelmäßigen Abstand der Beinpaare erkennbare Segmentierung zeigt sich in der regelmäßigen Aneinanderreihung von Hautöffnungen (Ostien), Ausscheidungsorganen (Nephridien) und Nervenzellkonzentrationen (Ganglien). Die einzelnen Körperabschnitte sind kaum spezialisiert; selbst der Kopf entwickelt sich nur wenig anders als ein beliebiges Hinterleibssegment. Die Segmentierung wird anscheinend durch das auch bei anderen Tiergruppen nachweisbare Gen engrailed festgelegt, das während der Embryonalentwicklung jeweils an der hinteren Segmentgrenze und in der Wachstumszone der Stummelfüße aktiviert wird.
Körperanhänge
 Stummelfüßer aus dem Amazonas-Regenwald in Peru
Stummelfüßer aus dem Amazonas-Regenwald in PeruDas namensgebende Merkmal der Stummelfüßer sind die fachlich als Lobopoden bezeichneten kegelförmigen, sackartigen Körperanhänge, die innen hohl sind und keine Gelenke aufweisen. Obwohl ihre Zahl zwischen den Arten beträchtlich variieren kann, sind sie sich im Aufbau grundsätzlich sehr ähnlich. Ihre Festigkeit beruht auf dem hydrostatischen Druck der enthaltenen Körperflüssigkeit.
Bewegt werden die Beine meist passiv durch Kontraktion und Dehnung des Gesamtkörpers. In jedem Bein verlaufen allerdings auch interne Muskeln, die es verkürzen und biegen können; aufgrund der fehlenden Gelenke kann diese Verbiegung an jeder Stelle entlang der Längsseite des Beines erfolgen.
Zwei verschiedene Organe befinden sich bei manchen Arten innerhalb der Beine:
- Cruraldrüsen sind am Beinansatz gelegene Drüsen, die bis in die Körperhöhle hineinreichen. Sie öffnen sich an den bauchseitig gelegenen Cruralpapillen, kleinen warzenförmigen Erhebungen, nach außen und sondern chemische Botenstoffe, so genannte Pheromone, ab. Ihr Name leitet sich von dem Lateinischen cruralis, „beinbezogen“, ab.
- Coxalvesikel, die auch als Coxalsäcke oder Coxalorgane bezeichnet werden, sind auf der bauchzugewandten Seite am Beinabsatz gelegene Säckchen, die nach außen gestülpt werden können und vermutlich der Wasseraufnahme dienen. Sie kommen nur innerhalb der Familie der Peripatidae vor und sind nach dem lateinischen Wort für „Hüfte“, coxa, benannt.
An den Füßen sitzt je ein Paar einziehbarer verhärteter (sklerotisierter) Krallen aus Chitin, von denen der wissenschaftliche Name des Taxons herrührt, der sich von dem Griechischen onyches, „Krallen“ und pherein, „tragen“, ableitet. Am Klauenansatz befinden sich drei bis sechs dornige „Kissen“, auf denen das Bein in Ruhestellung aufsetzt und auf welchen die Tiere auf glattem Untergrund laufen; die Krallen werden hauptsächlich auf unebenem Terrain eingesetzt, um festen Halt zu erlangen.
Neben den Beinpaaren gibt es drei weitere Körperanhänge, die sich an dem aus drei Segmenten bestehenden Kopf befinden:
- Am ersten Kopfsegment befindet sich ein Paar geringter Antennen, die der Sinneswahrnehmung dienen. Sie entsprechen wahrscheinlich nicht den Antennen der Gliederfüßer (Arthropoda), vielleicht aber deren "Lippen" oder Labrum. An ihrem Ansatz findet sich außer bei wenigen blinden Arten ein Paar einfacher Augen. Davor sind bei vielen australischen Arten diverse Grübchen, stilettartige Strukturen oder Dornen angelegt, deren Funktion noch nicht aufgeklärt ist, aber zumindest bei einigen Arten dem Transfer von Spermienpaketen (Spermatophoren) zu dienen scheint.
- Am zweiten Kopfsegment befindet sich bauchseitig die von sinnesempfindlichen "Lippen", dem Labrum, umgebene Mundöffnung; bei Stummelfüßern ist die Struktur allerdings ein muskulöser Auswuchs des Schlunds und daher trotz des gleichen Namens vermutlich nicht homolog zum Labrum der Gliederfüßer. Tief innerhalb der Mundhöhle liegen die scharfen, sichelförmigen Kiefer oder Mandibeln, die stark verhärtet sind und den Klauen der Beine ähneln, denen sie vermutlich homolog sind. Sie unterteilen sich in äußere und innere Mandibeln, sind mit feinen Zähnchen bedeckt und bewegen sich in Längsrichtung vor- und rückwärts; erlegte Beute kann so in feine Stücke zerraspelt werden.
- Am dritten Segment befinden sich links und rechts des Mundes zwei als Mundpapillen bezeichnete Öffnungen, in denen ein Paar großer, intern stark verästelter Schleimdrüsen mündet. Diese liegen etwa in der Mitte des Körpers und sondern einen dem Beutefang und der Verteidigung bestimmten milchigweißen Schleim ab. Manchmal ist der verbindende „Schleimleiter“ zu einem Reservoir verdickt, das den vorproduzierten Schleim zwischenspeichern kann. Schleimdrüsen sind wahrscheinlich modifizierte Cruraldrüsen.
Alle drei Strukturen entsprechen ihrer evolutionären Herkunft nach den Beinpaaren der anderen Segmente.
Haut und Muskulatur
Anders als die Gliederfüßer besitzen Stummelfüßer kein festes Außenskelett (Exoskelett). Stattdessen wird ihre Körperform wie bei einem wassergefüllten Ballon durch den leicht erhöhten Druck der Innenflüssigkeit auf die Körperwand aufrechterhalten: Weil diese Flüssigkeit bei den durch Muskelkraft auftretenden Drücken inkompressibel ist, also ihr Volumen nicht ändert, können Muskeln dagegen Arbeit verrichten. Wegen des Skelett-„Materials“ Wasser spricht man auch von einem Hydroskelett.
Die Körperwandung selbst besteht aus einer nicht-zelligen Außenhaut, der Cuticula, einer einlagigen Zellschicht, der Epidermis, als Innenhaut und in der Regel drei darunter gelegenen Muskelschichten, die in Bindegewebe eingebettet sind.
Die Cuticula ist etwa einen Mikrometer dünn, mit feinen Härchen besetzt und ähnelt in Zusammensetzung und Aufbau der Cuticula der Gliederfüßer. So besteht sie aus α-Chitin und diversen Proteinen, nicht jedoch aus Kollagen und lässt sich in eine außen liegende Epicuticula und eine darunter liegende Procuticula unterteilen, die sich ihrerseits aus Exo- und Endocuticula zusammensetzt. Dieser mehrschichtige Aufbau ist für die hohe Flexibilität der Außenhaut verantwortlich, die es den Tieren ermöglicht, sich noch in die engsten Spalten zu quetschen. Obwohl sie nach außen wasserabstoßend wirkt, ist sie nicht in der Lage, Feuchtigkeitsverluste zu verhindern, was zur Folge hat, dass Stummelfüßer nur in einem Mikroklima mit hoher Luftfeuchte leben können.
Die Oberfläche der Cuticula ist mit zahlreichen feinen Papillen übersät; die größeren von ihnen tragen die als Härchen sichtbaren Sinnesborsten. Die Papillen sind ihrerseits mit kleinen Schüppchen bedeckt und verleihen so der Haut ein samtartiges Aussehen. Sie fühlt sich bei Berührung auch entsprechend trocken-samtig an, wofür ihre wasserabweisende Natur verantwortlich ist. Im Rahmen der durch das Hormon Ecdyson vermittelten Häutung (Ecdysis) wird sie regelmäßig, manchmal alle vierzehn Tage, abgestoßen.
Erneuert wird sie dabei von der darunter gelegenen einschichtigen Innenhaut, der Epidermis, die im Gegensatz zur Cuticula aus lebenden Zellen besteht. Ihr unterliegt eine dicke Schicht Bindegewebe, das in erster Linie von Kollagenfasern gebildet wird, die entweder parallel oder senkrecht zur Körperlängsachse ausgerichtet sind. Innerhalb des Bindegewebes liegen drei ununterbrochene Schichten unspezialisierter glatter Muskulatur. Die äußere, verhältnismäßig dicke Schicht wird von den Ringmuskeln gebildet, die innerste von den ebenfalls voluminösen Längsmuskeln. Dazwischen liegen die dünnen Diagonalmuskeln, die sich schraubenförmig entlang der Körperachse nach vorne und hinten winden. Zwischen Ring- und Diagonalmuskulatur existieren feine Blutkanäle, die unterhalb der oberflächlich erkennbaren Querringe der Haut liegen und für diese Pseudosegmentierung genannte Musterung verantwortlich sind.
Unterhalb der innersten Muskelschicht liegt die Körperhöhle. Sie wird durch so genannte Dorsoventralmuskeln, die von der Mitte der Bauchseite quer durch sie hindurch zu den Rändern der Rückenseite verlaufen, im Querschnitt in drei Regionen geteilt: einen zentralen Mittelteil und zwei jeweils bauchseitig links und rechts gelegene Seitenteile, die auch die Beine einschließen.
Hämocoelom und Blutkreislauf
Die Körperhöhle selbst ist ein so genanntes Pseudocoelom, das im speziellen Fall auch als Hämocoelom bezeichnet wird. Ein Pseudocoel ist im Gegensatz zu einem echten Coelom nicht von einer echten, von embryonalem Mesoderm abgeleiteten Zellschicht umgeben. Ein Coelom ist jedoch um die Keimdrüsen und die der Ausscheidung dienenden Nephridien herum ausgebildet.
Wie der Name Hämocoelom bereits andeutet, ist die Körperhöhle mit einer blutartigen Flüssigkeit gefüllt, in die alle Organe eingebettet sind; auf diese Weise können sie leicht mit im Blut zirkulierenden Nährstoffen versorgt werden. Diese Flüssigkeit ist farblos, da sie keine Pigmente enthält; aus diesem Grund dient sie auch nur in geringem Maße dem Sauerstofftransport. In der Blutflüssigkeit bewegen sich zwei verschiedene Typen von Blutzellen oder Hämozyten, die Amöbozyten und Nephrozyten. Erstere haben vermutlich eine Funktion bei der Abwehr von Bakterien und anderen Fremdkörpern; bei einigen Arten spielen sie auch bei der Fortpflanzung eine Rolle. Nephrozyten speichern dagegen Giftstoffe oder wandeln sie in eine für die als Nephridien bezeichneten Ausscheidungsorgane verwertbare Form um.
Das Hämocoelom wird durch eine horizontale Scheidewand, das Diaphragma, in zwei Teile geteilt, den rückseitig gelegenen pericardialen Sinus und den bauchseitig gelegenen perivisceralen Sinus. Ersterer umgibt das schlauchförmige Herz, letzterer die anderen Organe. Das Diaphragma ist mehrfach perforiert, so dass ein Flüssigkeitsaustausch zwischen den beiden Teilhöhlungen möglich ist.
Das Herz selbst ist ein aus einschichtigem Epithelgewebe bestehender Ringmuskelschlauch mit je zwei seitlichen Öffnungen (Ostien) pro Segment. Während unbekannt ist, ob das Hinterende offen oder geschlossen ist, mündet es zur Vorderseite hin direkt in die Körperhöhlung. Da es abgesehen von den feinen, zwischen den Muskulaturschichten der Körperwandung befindlichen Blutgefäßen und einem Paar Arterien, welche die Antennen versorgen, keine Adern gibt, spricht man von einem offenen Blutkreislauf.
Der zeitliche Ablauf des Pumpvorgangs lässt sich in zwei Teile zerlegen, Diastole und Systole. Während der Diastole fließt das Blut aus dem pericardialen (herznahen) Sinus durch die Ostien in das Herz. Sobald die Systole beginnt, schließen sich die Ostien, die Herzmuskulatur zieht sich zusammen, verringert so das Herzvolumen und pumpt auf diese Weise Blut aus dem Vorderende des Herzens in das periviscerale (organnahe) Hämocoelom. Die verschiedenen Gewebe dort werden auf diese Weise mit Nährstoffen versorgt, bevor das Blut schließlich wieder durch die Perforationen des Diaphragmas in den pericardialen Sinus zurückgelangt. Neben der Pumptätigkeit des Herzens haben auch Körperbewegungen einen Einfluss auf die Blutzirkulation.
Atmung
Die Sauerstoffaufnahme der Tiere erfolgt zum einen durch einfache Diffusion über die gesamte Körperoberfläche hinweg, bei einigen Arten möglicherweise auch über ihre Coxalvesikel an den Stummelbeinen. Am wichtigsten ist jedoch der Gasaustausch über feine unverzweigte Röhrchen, die Tracheen, die von der Körperoberfläche tief in den Körper zu den verschiedenen Organen, ganz besonders aber zum Herz hinziehen.
Die Wandung dieser meist weniger als drei Mikrometer dicken Strukturen besteht nur aus einem extrem dünnen Häutchen, durch die der Sauerstoff leicht diffundieren kann. Sie entspringen an winzigen Öffnungen, den Spirakeln, die ihrerseits jeweils zu mehreren in einer dellenförmigen Einbuchtung der Außenhaut, dem Atrium, zusammengefasst sind. Die Zahl der so entstehenden Tracheenbündel lässt sich mit durchschnittlich 75 je Körpersegment angeben; gehäuft treten sie vor allem auf der Rückseite der Tiere auf.
Anders als Gliederfüßer, die ihre Tracheenöffnungen kontrollieren können, sind die Stummelfüßer dazu nicht in der Lage. Ihre Tracheen sind immer geöffnet, was bei Trockenheit große Wasserverluste zur Folge hat. Aus diesem Grund sind Stummelfüßer auf Lebensräume mit hoher Luftfeuchtigkeit angewiesen.
Verdauungssystem
Der Verdauungstrakt beginnt mit dem subterminal, also etwas hinter dem vordersten Punkt des Körpers auf der Bauchseite gelegenen Mund, in dem Beute durch die mit feinen Zähnchen besetzten Kiefer mechanisch zerkleinert werden kann. Zwei Speicheldrüsen münden durch einen gemeinsamen Leiter in den sich anschließenden Schlund, der bereits den ersten Teil des Vorderdarms darstellt. Der von ihnen produzierte Speichel enthält Schleim und hydrolytische Enzyme, die in und außerhalb der Mundhöhle bereits die Verdauung einleiten. Evolutionsgeschichtlich leiten sich die Speicheldrüsen wahrscheinlich von als Nephridien bezeichneten Ausscheidungsorganen ab, die sich als homologe Organe in den anderen Körpersegmenten befinden.
Der Schlund selbst ist sehr muskulös und dient dazu, die vorverdaute teilverflüssigte Nahrung einzusaugen und durch die Speiseröhre, die den hinteren Teil des Vorderdarms bildet, in den Mitteldarm zu pumpen. Dieser ist anders als der Vorderdarm nicht mit Cuticula ausgekleidet und besteht nur aus einer einlagigen Schicht Epithelgewebe, die anders als bei anderen Tieren keine auffälligen Einbuchtungen aufweist. Beim Eintritt in den Mitteldarm werden Nahrungspartikel von einer schleimhaltigen peritrophen Membran umhüllt, die eine mechanische Beschädigung des Darmgewebes durch scharfkantige Partikel verhindern soll. Das Darmepithel selbst sondert weitere Verdauungsenzyme ab und absorbiert die aufgeschlossenen Nährstoffe; der Großteil der Verdauung findet allerdings bereits extern oder in der Mundhöhle statt. Unverdauliche Reste gelangen in den auch Rektum genannten Hinterdarm, der wiederum mit Cuticula ausgekleidet ist und sich bauchseitig nahe dem Hinterende der Tiere in den After öffnet.
Ausscheidungsorgane
In fast jedem Segment befindet sich ein Paar als Nephridien bezeichneter Ausscheidungsorgane, die sich von Coelomgewebe ableiten. Sie bestehen jeweils aus einem kleinen Säckchen, das über einen geißelbesetzten Gang, Nephridiodukt genannt, mit einer bauchseitig am nächstgelegenen Beinansatz befindlichen Öffnung, der Nephridiopore, verbunden ist. Das Säckchen ist mit speziellen Zellen, den Podozyten besetzt, die eine Ultrafiltration des Blutes durch die Trennwand von Hämocoelom und Nephridium ermöglichen. Der Primärurin wird durch selektive Rückgewinnung von Nährstoffen und Wasser sowie durch Absonderung von Gift- und Abfallstoffen im Nephridiodukt noch in seiner Zusammensetzung verändert, bevor er an der Nephridiopore an die Außenwelt abgegeben wird. Das wichtigste stickstoffhaltige Ausscheidungsprodukt ist die wasserunlösliche Harnsäure; sie kann in festem Zustand mit nur wenig Wasser an die Umgebung abgegeben werden. Dieser als uricotelisch bezeichnete Ausscheidungsmodus stellt eine Anpassung an das Landleben und die Notwendigkeit, sparsam mit Wasser umzugehen, dar.
Ein ehemaliges Nephridienpaar im Kopf wurde sekundär in die Speicheldrüsen umgewandelt, ein weiteres im letzten Segment der Männchen übernimmt heute eine Drüsenfunktion, die anscheinend bei der Fortpflanzung zum Einsatz kommt.
Nervensystem
Das Nervensystem besteht in erster Linie aus dem Gehirn und zwei bauchseitigen Nervensträngen, die durch zahlreiche Querverbindungen miteinander verbunden sind, so dass man auch von einem „Strickleiternervensystem“ spricht.
Das vorne und rückseitig gelegene Gehirn besteht aus einer linken und einer rechten Hälfte, die zusammen mit Nervenverbindungen einen Ring um die Speiseröhre bilden. Eine Dreiteilung des Gehirns in Richtung der Körperlängsachse, wie man sie bei den eng verwandten Gliederfüßern (Arthropoda) findet, lässt sich nur im embryonalen Zustand beobachten.
Vom Gehirn gehen etwa fünfzehn Nervenpaare aus, die sowohl sensorische, als auch motorische Nervenzellen enthalten. Die Hauptnerven des Kopfbereichs sind die optischen Nerven, die Augen und Gehirn verbinden und zwei Sinnesnerven, die in die Antennen ziehen und für die Vermittlung von Berührungs- und chemischen Reizen verantwortlich sind. Die wichtigsten Körpernerven sind dagegen zwei weit auseinander liegende, bauchseitige Nervenstränge, die sich den gesamten Körper entlang längsseitig nach hinten ziehen. Pro Segment findet sich je ein Paar unauffälliger Nervenzellkonzentrationen, die Ganglien, die jeweils durch neun oder zehn querlaufende Nerven miteinander verbunden sind. Körperwand und -anhänge werden durch gemischte Nerven versorgt, die sowohl sensorische, der Sinnenswahrnehmung dienende als auch motorische, die Muskeln ansprechende Signale weiterleiten.
Sinnesorgane
Der gesamte Körper inklusive der Stummelfüße ist mit zahlreichen Papillen übersät, warzigen Vorsprüngen, die eine auf mechanische Reize ansprechende (mechanorezeptive) Borste an der Spitze tragen, die jeweils mit weiter innen liegenden Sinnesnervenzellen verbunden ist. Auch die Mundpapillen, die Ausgänge der Schleimdrüsen, haben vermutlich eine Funktion bei der Sinneswahrnehmung.
Die auf den "Lippen", dem Labrum, gelegenen Sinneszellen oder Sensillen dienen dagegen der Wahrnehmung chemischer Reize, weshalb man von Chemorezeptoren spricht. Sie finden sich auch auf den beiden Antennen, die als die wichtigsten Sinnesorgane der Stummelfüßer angesehen werden können.
Mit Ausnahme einiger weniger, meist höhlenlebender Arten befindet sich am Ansatz der Antennen je ein einfach gebautes Auge (Ocellus), das aus einer chitinhaltigen Kugellinse sowie Hornhaut und Netzhaut besteht und durch je einen optischen Nerv mit dem Gehirn verbunden ist. Die Netzhaut setzt sich aus zahlreichen Pigmentzellen und Photorezeptoren zusammen; bei letzteren handelt es sich um leicht modifizierte Geißelzellen, deren Geißelmembran ein lichtempfindliches Pigment auf der Oberfläche trägt.
Fortpflanzungsorgane
Beide Geschlechter verfügen über paarige Keimdrüsen, die durch als Gonodukte bezeichnete Ausführgänge in eine gemeinsame Geschlechtsöffnung, die Gonopore, münden, welche sich auf der hinteren Bauchseite befindet. Sowohl Keimdrüsen als auch Gonodukte leiten sich von echtem Coelomgewebe ab.
Bei den Weibchen sind die beiden Eierstöcke in der Mitte zusammengewachsen und an die horizontal verlaufende Scheidewand geheftet. Die Gonodukte sind unterschiedlich ausgeprägt, je nachdem, ob es sich um lebendgebärende oder eierlegende Arten handelt. Bei Ersteren teilt sich jeder Ausführgang in einen schlanken Eileiter und eine geräumige „Gebärmutter“, den Uterus, in dem die Embryos heranwachsen. Die nur einfach vorhandene Vagina, mit der beide Uteri verbunden sind, mündet an der Gonopore nach außen. Bei eierlegenden Arten, deren Gonodukte gleichförmig aufgebaut sind, liegt die Geschlechtsöffnung an der Spitze eines langen Eierlegeapparats, des Ovipositors. Die Weibchen vieler Arten verfügen darüber hinaus über einen Receptaculum seminis genannten Samenspeicher, in dem Spermazellen der Männchen vorübergehend oder auch längerfristig aufbewahrt werden können.
Männchen verfügen über zwei getrennte Hoden sowie die dazugehörigen Samenbläschen (vesicula seminalis) und Ausführgänge (vasa efferentia). Letztere vereinigen sich zu einem gemeinsamen Samenleiter, dem vas deferens, der sich wiederum zum Ejakulationsleiter verbreitert an der Gonopore nach außen öffnet. Direkt neben beziehungsweise hinter dieser liegen bei den Männchen zwei Paar spezieller Drüsen, die vermutlich eine Hilfsfunktion bei der Fortpflanzung haben; die Drüsen des hinteren Paars werden auch als Analdrüsen bezeichnet.
Eine penisartige Struktur konnte bisher nur bei Männchen der Gattung Paraperipatus nachgewiesen, aber noch nicht in Aktion beobachtet werden. Männchen zahlreicher australischer Arten weisen dagegen wie bereits erwähnt spezielle Strukturen am Kopf auf, die anscheinend Aufgaben bei der Samenübertragung auf die Weibchen übernehmen.
Verbreitung und Lebensraum
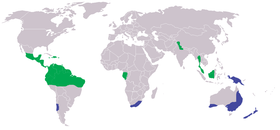 Verbreitung der Onychophora: Peripatidae (grün), Peripatopsidae (blau)
Verbreitung der Onychophora: Peripatidae (grün), Peripatopsidae (blau)Stummelfüßer leben in tropischen Lebensräumen und in der gemäßigten Zone der Südhalbkugel; sie weisen sowohl eine zirkumtropische, als auch eine zirkumaustrale Verbreitung auf. Einzelne Arten findet man in Mittel- und Südamerika, auf den Karibischen Inseln, im äquatorialen Westafrika und in Südafrika, des Weiteren in Nordindien, Indonesien und Teilen Malaysias, auf Neu-Guinea, in Australien und in Neuseeland. Die Gesamtheit der heutigen Verbreitungsgebiete weist auf den wahrscheinlichen Ursprung des Taxons auf dem ehemaligen Superkontinent Gondwana hin, weswegen man auch von einer Gondwana-Verteilung spricht.
Alle Stummelfüßer sind landlebend (terrestrisch) und bevorzugen dunkle Lebensräume mit hoher Luftfeuchtigkeit. In Frage kommen dafür insbesondere die Regenwälder der Tropen und gemäßigten Zonen, wo sich die Tiere in Mooskissen, in der Laubstreu des Bodens, unter Baumstämmen und Steinen, in verrottendem Holz oder in Termitengängen finden lassen. Auch in unbewaldetem Grasland kommen sie vor, wenn ausreichend Bodenspalten existieren, in die sie sich während des Tages zurückziehen können.
Zwei Arten leben in Höhlen, einem Lebensraum, an den sie durch ihre Fähigkeit, sich in kleinste Ritzen zwängen zu können, hervorragend angepasst sind und der ihnen konstante Lebensbedingungen garantiert. Da die entscheidenden Voraussetzungen für ein Höhlenleben wahrscheinlich bereits vor der Besiedlung dieses Habitats vorlagen, spricht man auch von Exaptation. Die Landwirtschaft hat Stummelfüßern anscheinend neue Lebensräume erschlossen; jedenfalls finden sie sich in Zentralamerika und der Karibik auch in künstlich angelegten Kakao- und Bananenplantagen.
Weil die Austrocknungsgefahr tagsüber und bei trockenem Wetter am größten ist, verwundert es nicht, dass Stummelfüßer in der Regel nachts und bei Regenwetter aktiv sind. Unter kalten oder trockenen Umweltbedingungen suchen sie aktiv Bodenspalten auf, in denen sie ihren Körper in einen Ruhezustand versetzen. Von starken Lichtquellen wenden sie sich ab, weswegen man sie auch als negativ phototaktisch bezeichnet.
Die bisher größte gemessene Populationsdichte liegt mit etwa 10 Individuen pro Quadratmeter Boden sehr niedrig; Stummelfüßer sind auch in ihren Lebensräumen nur sehr schwer zu finden.
Fortbewegung
Um von der Stelle zu kommen, krabbeln die Tiere mithilfe ihrer Beine vorwärts; sie bewegen dabei anders als die Gliederfüßer immer beide Beine eines Paares gleichzeitig. Eine Berührung der Bauchseite mit dem Substrat wird nach Möglichkeit vermieden, der Körper also durch die Stummelfüße oberhalb des Bodens gehalten. Die an den Füßen befindlichen Klauen werden nur auf hartem, rauem Untergrund eingesetzt, wo ein fester Halt erforderlich ist; auf weichen Substraten wie etwa Moos laufen die Tiere auf den am Klauenansatz befindlichen Fußkissen.
Die eigentliche Fortbewegung geschieht weniger durch den Einsatz der Beinmuskulatur, sondern stattdessen durch eine lokale Veränderung der Körperlänge. Diese kann von den Tieren durch Einsatz der Ring- und Längsmuskulatur beeinflusst werden: Wird erstere kontrahiert, verringert sich der Körperquerschnitt und das entsprechende Segment dehnt sich, da sein Volumen aufgrund der enthaltenen inkompressiblen Flüssigkeit des Hämocoeloms konstant bleiben muss - dies ist die übliche Wirkungsweise eines Hydroskeletts wie es auch bei verschiedenen Würmern zum Einsatz kommt. Durch die Dehnung heben sich die Beine des betroffenen Körperabschnitts und schwingen vorwärts. Lokale Kontraktion der Längsmuskeln verkürzt dagegen das entsprechende Segment und bewegt auf diese Weise das zugehörige Beinpaar, dass sich nun im Kontakt mit dem Substrat befindet, nach hinten; dieser Teil des Bewegungszyklus ist der effektive Beinschlag, der für die eigentliche Fortbewegung verantwortlich ist. Durch das Nervensystem werden die einzelnen Muskeldehnungen und -entspannungen der Segmente derart aufeinander abgestimmt, dass Kontraktionswellen durch den Körper laufen, die dazu führen, dass ein Beinpaar nach dem anderen nach vorne schwingt und sich dann auf dem Untergrund nach hinten bewegt. Die auf diese Weise erzielten Geschwindigkeiten schwanken zwischen etwa einem Millimeter und etwas mehr als einem Zentimeter pro Sekunde.
Ernährung
Stummelfüßer sind räuberische Organismen, die in der Lage sind, Tiere zu erbeuten, die erheblich größer sind als sie selbst. Zum Beutespektrum gehören etwa Asseln (Isopoda), Termiten (Isoptera), Echte Grillen (Gryllidae), Staubläuse (Psocoptera), Schaben (Blattodea), Tausendfüßer (Myriapoda), Spinnen (Araneae), diverse Würmer und selbst größere Schnecken (Gastropoda). Sie gelten als ökologisches Äquivalent der Hundertfüßer (Chilopoda).
Potentielle Opfer spüren sie zumeist mit Hilfe der Antennen auf und verfolgen sie dann bis in kleinste Ritzen. Während kleinere Beute direkt getötet wird, nutzen sie bei größeren Tieren eine weiße leimartige und stark proteinhaltige Flüssigkeit, die von den beiden Schleimdrüsen produziert wird, um ihre Opfer unbeweglich zu machen. Sie wird durch die Poren der Mundpapillen über eine Entfernung von bis zu 30 Zentimetern verspritzt und härtet an Luft sehr schnell aus, so dass sich die Beute in der stark klebrigen Substanz verfängt und schließlich unbeweglich wird. Auf der wasserabweisenden Haut der Stummelfüßer selbst klebt die Substanz dagegen nicht, so dass die Tiere sich ihrem Opfer ungefährdet auf kürzeste Distanz nähern können. Durch Injektion toxischen Speichels wird es nun abgetötet und vorverdaut; die scharfen Kiefer zerschneiden die Beute dann in feine Stücke, die über den Mund in den Verdauungstrakt gelangen.
Die räuberische Lebensweise ist vermutlich ein Ergebnis der hohen Feuchtebedürftigkeit der Organismen: Wegen der beständigen Austrocknungsgefahr stehen oft nur wenige Stunden am Tag zur Nahrungsbeschaffung zur Verfügung, so dass eine starke Selektion für ein niedriges Kosten-Nutzen-Verhältnis besteht, das sich kaum durch eine pflanzliche Ernährung erreichen lässt.
Fressfeinde und Parasiten
Wichtige Fressfeinde der Tiere sind in erster Linie diverse Spinnen (Araneae) und Hundertfüßer (Chilopoda), daneben aber auch Nagetiere (Rodentia) und Vögel, in Zentralamerika etwa die Gilbdrossel (Turdus grayi). Die zu den Korallenottern (Micrurus) gezählte Schlangenart Micrurus hemprichii ernährt sich sogar beinahe ausschließlich von Stummelfüßern. Zur Abwehr rollen sich manche Arten reflexartig zu einer Spirale zusammen; daneben können sich die Tiere auch durch Schleimauswurf gegen kleinere Gegner zur Wehr setzen.
Diverse Milben (Acari) sind als Ectoparasiten bekannt, die sich auf der Haut der Stummelfüßer festsetzen. Bei Hautverletzungen kommt es zudem meist zu bakteriellen Infektionen, die fast immer tödlich enden.
Fortpflanzung und Lebenszyklus
Mit Ausnahme einer einzigen Art, Epiperipatus inthurni, bei der bisher keine Männchen nachgewiesen werden konnten und die sich nachweislich parthenogenetisch, also ohne Befruchtungsvorgang, vermehrt, findet die Fortpflanzung bei Stummelfüßern immer auf sexuellem Wege statt.
Alle Tiere sind grundsätzlich getrenntgeschlechtlich und weisen in vielen Fällen einen merklichen Geschlechtsdimorphismus auf - die Weibchen sind meist größer als die Männchen und verfügen, falls die Beinzahl innerhalb einer Art variabel ist, auch über mehr Beine. Bei vielen Arten werden sie nur einmal im Leben befruchtet, was dazu geführt hat, dass die Begattung manchmal schon stattfindet, wenn die Fortpflanzungsorgane der Weibchen noch nicht ausgereift sind, bei Macroperipatus torquatus beispielsweise schon im Alter von drei Monaten. Die übertragenen Spermien werden in diesem Fall in einem speziellen Speicherorgan aufbewahrt, wo sie für längere Zeiträume befruchtungsfähig bleiben können.
Die Befruchtung findet bei Stummelfüßern grundsätzlich intern statt, die Art der Samenübertragung variiert allerdings recht stark. Bei den meisten Arten, etwa denen der Gattung Peripatus, wird ein Spermienpaket, die Spermatophore, in der Geschlechtsöffnung des Weibchens platziert. Wie im Einzelnen dies geschieht, ist in den meisten Fällen allerdings noch unbekannt, ein regelrechter Penis ist nur bei Arten der Gattung Paraperipatus nachgewiesen. Bei zahlreichen australischen Arten existieren Grübchen oder spezielle stilett- oder hakenförmige Strukturen am Kopf; bei Florelliceps stutchburyae etwa presst das Männchen einen langen Dorn gegen die Geschlechtsöffnung der Weibchen und positioniert sehr wahrscheinlich auf diese Weise seine Spermatophore dort. Es wird dabei durch das Weibchen unterstützt, das mit den Klauen des letzten Beinpaares das Männchen umklammert hält. Besonders ausgefallen fällt die Spermienübertragung bei zwei Arten der Gattung Peripatopsis aus. Hier platziert das Männchen zwei millimetergroße Spermatophoren auf dem Rücken oder den Flanken des Weibchens. Amöbozyten aus dem Blut des Weibchens sammeln sich daraufhin auf der Innenseite der Ablagerungsstelle und bauen durch Sekretion von Enzymen die unter dem Spermienpaket befindliche Körperwandung sowie die Hülle der Spermatophore ab. Die dadurch freigesetzten Spermien bewegen sich dann frei durch das Hämocoel zu den Eierstöcken, durchdringen deren Außenwand und befruchten schließlich die Eizellen. Warum es durch die selbstverursachte Hautverletzung nicht zu bakteriellen Infektionen kommt, ist noch unverstanden.
Bei Stummelfüßern findet man sowohl eierlegende (ovipare), eierlebendgebärende (ovovivipare) und lebendgebärende (vivipare) Formen.
- Erstere finden sich nur in der Familie der Peripatopsidae, häufig in Regionen mit unstetem Nahrungsangebot oder wechselhaftem Klima. Die etwa 1,3 bis 2 Millimeter großen, stark dotterhaltigen Eier sind in diesem Fall von einer schützenden Eierschale aus Chitin umhüllt; mütterliche Fürsorge ist unbekannt.
- Die Mehrzahl der Arten ist eierlebendgebärend: Die mittelgroßen, lediglich von einer Membran umhüllten Eier verbleiben in der „Gebärmutter“, dem Uterus, die Embryos ernähren sich aber von der moderaten Menge Dotter, die in den Eiern enthalten ist und werden nicht gesondert von der Mutter versorgt, man bezeichnet sie daher auch als lecitotroph; sie schlüpfen erst kurz vor der Geburt. Vermutlich stellt dies den ursprünglichen Fortpflanzungsmodus der Stummelfüßer dar, das heißt sowohl ovipare als auch vivipare Arten haben sich aus den ovoviviparen Formen entwickelt.
- Echt lebendgebärende Arten finden sich in beiden Familien, besonders in tropischen Regionen mit einem über das Jahr hinweg stabilen Klima und gleichmäßigem Nahrungsangebot. Die sich aus den nur mikrometergroßen Eiern entwickelnden Embryonen werden im Uterus von ihrer Mutter ernährt; sie werden daher als matrotroph bezeichnet. Die Versorgung erfolgt entweder durch mütterliche Sekretabgabe in den Uterus oder durch eine echte Gewebeverbindung zwischen dem Gebärmutterepithel und dem heranwachsenden Embryo, eine so genannte Plazenta. Ersteres kommt nur außerhalb des amerikanischen Kontinents vor, letzteres findet sich in erster Linie in Amerika und der Karibik, seltener in der alten Welt. Die Tragzeit, an deren Ende die Jungtiere bereits weitgehend entwickelt geboren werden, kann bis zu 15 Monate betragen. Die im Uterus eines Weibchens vorfindlichen Embryonen müssen nicht notwendigerweise gleichaltrig sein; es kann durchaus vorkommen, dass sich dort zahlreiche unterschiedlich entwickelte Tiere finden, die zudem von verschiedenen Männchen abstammen.
Die Zahl der Nachkommen eines Weibchens schwankt zwischen 1 und 23 pro Jahr; die zwischen 6 und 17 Monate andauernde Entwicklung von der befruchteten Eizelle zum Erwachsenenstadium verläuft grundsätzlich direkt ohne Larvenstadium; vermutlich ist dies auch der ursprüngliche Entwicklungsmodus. Maximal können Stummelfüßer bis zu 6 Jahre alt werden.
Gefährdung
Die globale Gefährdung der Stummelfüßer-Vielfalt lässt sich nur schwer einschätzen; viele Arten sind nur von der Typ-Lokalität, also dem Ort, von dem das erste beschriebene Exemplar stammte, bekannt. Niedrige Populationsdichten, die meist nächtliche Lebensweise der Tiere, möglicherweise noch unerfasste jahreszeitliche Einflüsse und geschlechtsspezifische Unterschiede erschweren die Erhebung vertrauenswürdiger Angaben zusätzlich.
Bisher wurden nur elf Arten hinreichend genau untersucht, um Angaben über ihre Häufigkeit machen zu können, davon gelten drei, Opistopatus roseus, Speleoperipatus spelaeus und Peripatopsis leonina, als kritisch bedroht, letztere Art ist vermutlich sogar bereits ausgestorben. Zwei weitere, Macroperipatus insularis und Tasmanipatus anophtalmus werden von der International Union for the Conservation of Nature and Natural Resources als bedroht eingeschätzt, vier weitere Arten gelten als gefährdet.
Ursache der Bedrohung ist in erster Linie der Verlust und die Fragmentierung des Lebensraums durch zunehmende Industrialisierung, Trockenlegung von Feuchtgebieten und Brandrodungen zu Landwirtschaftszwecken. Viele Arten haben dazu eine von Natur aus niedrige Populationsdichte und ein geografisch eng beschränktes Verbreitungsgebiet, was dazu führt, dass schon kleinere lokale Störungen der betroffenen Ökosysteme zum Aussterben ganzer Populationen und gegebenenfalls Arten führen können. Regional spielt daneben auch das Sammeln von Tieren für Universitäten oder Forschungsinstitute eine Rolle.
Der Schutz der Stummelfüßer ist regional sehr unterschiedlich ausgeprägt; in manchen Staaten wie Südafrika existieren Sammelbeschränkungen und Exportkontrollen, in anderen wie Australien nur letztere; aus vielen Staaten sind überhaupt keine spezifischen Schutzmaßnahmen bekannt. Weltweit einzigartig ist ein Schutzprogramm in Tasmanien: In einem dortigen Forstbezirk kommt ein eigener „velvet worm conservation plan“ zum Einsatz, der speziell auf eine Stummelfüßer-Art zugeschnitten ist.
Gezüchtet werden können Stummelfüßer bisher noch nicht. Entsprechende Überlegungen bestehen aber, nicht nur mit dem Hintergedanken, Populationen für spätere Auswilderungen bereitstellen zu können, sondern auch, um die betroffene Öffentlichkeit besser über die Tiere und die Notwendigkeit ihres Schutzes aufzuklären. Aufgrund der oft bunten Farben und der großen Bedeutung für die Evolutionsgeschichte käme zum Beispiel eine Haltung in „Insektenzoos“ in Frage.
Stammesgeschichte
Moderne Formen
Unter den heutigen Formen sind die Stummelfüßer wahrscheinlich sehr eng mit den Gliederfüßern (Arthropoda) verwandt, einem sehr umfangreichen Taxon, zu dem man etwa die Krebstiere (Crustacea), Insekten (Insecta) und Spinnentiere (Arachnida) zählt. Mit ihnen teilen sie unter anderem das gehäutete und aus α-Chitin sowie nicht-kollagenen Proteinen zusammengesetzte Außenskelett, die von echtem Coelomgewebe umgebenen Keimdrüsen und Ausscheidungsorgane, das offene Blutsystem mit rückseitig gelegenem Schlauchherzen, eine in pericardiale und periviscerale Höhlen aufgeteilte Leibeshöhle, Atmung durch Tracheen und eine ähnliche Embryonalentwicklung. Dazu kommt die Segmentierung mit je zwei Körperanhängen pro Segment. Antennen, Mandibeln und Oralpapillen sind dagegen vermutlich nicht zu den entsprechenden Körperteilen der Gliederfüßer homolog, das heißt, sie haben sich wohl unabhängig von diesen entwickelt.
Eine weitere eng verwandte Gruppe sind die relativ obskuren Bärtierchen (Tardigrada), denen allerdings aufgrund ihrer sehr geringen Größe eine Reihe von Merkmalen der Stummel- und Gliederfüßer wie Blutkreislauf und separate Atmungsstrukturen fehlen. Gemeinsam bilden Stummelfüßer, Gliederfüßer und Bärtierchen ein monophyletisches Taxon, die Panarthropoda, das heißt, alle drei Gruppen zusammengenommen umfassen alle Nachkommen ihres letzten gemeinsamen Vorfahrens.
Innerhalb der Panarthropoda wurden die Stummelfüßer meist aufgrund einer gewissen Formverwandtschaft mit den Bärtierchen zu einem Taxon Protoarthropoda zusammengefasst. Mit dieser Bezeichnung sollte zum Ausdruck gebracht werden, dass beide, Stummelfüßer und Bärtierchen, noch nicht die volle Entwicklungshöhe der Gliederfüßer erreicht haben. Moderne systematische Theorien verwerfen derartige Vorstellungen von „primitiven“ und „hoch entwickelten“ Lebensformen und orientieren sich stattdessen ausschließlich an den stammesgeschichtlichen Verwandtschaftsverhältnissen der beteiligten Taxa. Diese sind noch nicht zur Gänze aufgeklärt; es gilt jedoch als relativ wahrscheinlich, dass die Schwestergruppe der Stummelfüßer von einem Taxon gebildet wird, das sowohl Bärtierchen und Gliederfüßer umfasst und als Tactopoda (manchmal aber auch Tritocerebra) bezeichnet wird:
Panarthropoda |--Stummelfüßer |--Tactopoda |--Bärtierchen (Tardigrada) |--Gliederfüßer (Arthropoda)
In die weitere Verwandtschaft der Stummelfüßer wurden lange Zeit die Ringelwürmer (Annelida) gestellt. Mit ihnen verbindet sie unter anderem der wurmartige Körper, die dünne und flexible Außenhaut, eine schichtige Muskulatur, paarig angelegte Ausscheidungsorgane sowie ein einfach gebautes Gehirn und einfache Augen. Ausschlaggebend war jedoch die bei beiden Gruppen vorhandene Segmentierung, die nur wenig Spezialisierung der Einzelsegmente erkennen lässt. Die Parapodien genannten Körperanhänge der Ringelwürmer entsprechen demnach den Gliedmaßen der Stummelfüßer.
In dem von Georges Cuvier geprägten Articulata-Konzept gelten die Stummelfüßer daher als evolutionäres Bindeglied zwischen Ringelwürmern und Gliederfüßern: Wurmartige Vorläufer entwickelten demnach zuerst Parapodien, die sich dann über Stummelbeine als Zwischenglied letztlich zu den Körperanhängen der Gliederfüßer weiterentwickelten. Die Stummelfüßer selbst galten aufgrund ihres strukturellen Konservatismus folglich als lebende Fossilien. Diese Ansicht kommt paradigmatisch in der Aussage des französischen Zoologen A. Vandel zum Ausdruck:
- Onychophorans can be considered highly evolved annelids, adapted to terrestrial life, which announced prophetically the Arthropoda. They are a lateral branch which has endured from ancient times until today, without important modifications.
- Onychophoren können als hoch entwickelte und an das Landleben angepasste Ringelwürmer angesehen werden, die prophetisch das Erscheinen der Gliederfüßer ankündigten. Sie sind ein Seitenast, der ohne relevante Veränderungen aus Urzeiten bis heute überdauert hat.1
Die moderne Systematik bemüht sich, Kriterien wie eine "höhere" oder „niedrigere“ Entwicklungsstufe der betrachteten Lebewesen ebenso außen vor zu lassen wie Unterscheidungen in Haupt- und Seitenäste - ihr gelten nur die aufgrund kladistischer Methodik erschlossenen Verwandtschaftsbeziehungen als real. Für das Articulata-Konzept spricht aus dieser Sicht immer noch der bei allen betroffenen Tiergruppen segmentierte Körper, das Vorhandensein gepaarter Segmentanhänge, die paarweise Anlage von Ausscheidungsorganen in jedem Segment und vor allem ein strickleiterartig aufgebautes Nervensystem auf der Basis eines bauchseitig gelegenen Doppelnervenstrangs.
Die Vertreter eines alternativen und heute mehrheitlich bevorzugten Konzepts, der so genannten Ecdysozoa-Hypothese, stellen dagegen Ringelwürmer und Panarthropoda in zwei sehr unterschiedliche Großgruppen, die Lophotrochozoen (Lophotrochozoa) einerseits und die Häutungstiere (Ecdysozoa) andererseits.
Sie gehen damit davon aus, dass es sich bei den vorgenannten Gemeinsamkeiten von Ringelwürmern und Stummelfüßern entweder um konvergent entstandene oder um „primitive“ Merkmale, so genannte Symplesiomorphien, handelt, die beide betroffene Gruppen unverändert von den gemeinsamen Vorfahren von Lophotrochozoen und Häutungstieren übernommen haben. Im ersteren Falle hätte sich etwa das Strickleitersystem im Laufe der Evolutionsgeschichte zweimal unabhängig entwickelt, im zweiteren handelte es sich um eine sehr altes Merkmal, das keine besonders enge Verwandtschaft zwischen Ringelwürmern und Panarthropoda begründen kann.
Das Ecdysozoa-Konzept sieht die weitere Verwandtschaft der Stummelfüßer in einem Cycloneuralia genannten Taxon, zu dem man neben Faden- (Nematoda) und Saitenwürmern (Nematomorpha) auch drei eher obskure Gruppen, Hakenrüssler (Kinorhyncha), Priapswürmer (Priapulida) und Korsetttierchen (Loricifera) zählt.
Urmünder (Protostomia) |--Häutungstiere (Ecdysozoa) | |--Panarthropoda (Gliederfüßer, Stummelfüßer, Bärtierchen) | |--Cycloneuralia (Fadenwürmer, Saitenwürmer und weitere) |--Lophotrochozoen (Lophotrochozoa) (Ringelwürmer, Weichtiere und weitere) |--weitere
Besonders charakteristisch für die Cycloneuralia ist ein um die Mundöffnung herumgelegener, zirkumoraler, Nervenring, den die Befürworter des Ecdysozoa-Konzepts in modifizierter Form auch im Innervierungsmuster, also den Details der Nervenanordnung, der Panarthropoda wiedererkennen. Hinzu kommen ein allen Gruppen gemeinsamer Häutungsmechanismus, die Ecdysis, und molekularbiologisch aufgefundene Gemeinsamkeiten. Ein Problem der Ecdysozoa-Hypothese ist die subterminale Mundstellung der Stummelfüßer: Anders als bei den Cycloneuralia befindet sich der Mund nicht am Vorderende des Körpers, sondern liegt bauchseitig dahinter. Es gibt allerdings durch entwicklungsbiologische Untersuchungen, insbesondere hinsichtlich der Entstehung der Kopfnerven, Hinweise darauf, dass dies nicht immer der Fall war und der Mund ursprünglich terminal, also endständig angelegt war. Dies wird durch den Fossilbefund gestützt.
Ausgestorbene Formen
Die fossile Überlieferung der Stummelfüßer schließt Fossilien aus drei verschiedenen Zeitabschnitten ein, namentlich den frühen Epochen des Erdaltertums Kambrium und Ordovizium, dem späteren Karbon sowie dem Tertiär in der Erdneuzeit. Die kambrischen und ordovizischen Formen, deren Zugehörigkeit zu den Stummelfüßern umstritten ist, lebten noch allesamt im Meer, während die späteren Arten bereits als terrestrisch gelten.
Traditionell werden alle fossilen Formen in ein separates Taxon Xenusia gestellt, während die modernen als Euonychophora bezeichnet werden. Diese Klassifikation sagt allerdings nichts über die tatsächlichen stammesgeschichtlichen Zusammenhänge aus, weshalb sie bei kladistisch arbeitenden Taxonomen nicht akzeptiert ist.
Lobopoden
Die ältesten annähernd vollständig erhaltenen Fossilien stammen aus dem unteren Kambrium. Die auffälligsten Merkmale, die diese als Lobopoden bezeichneten Formen mit den modernen Stummelfüßern teilen, sind die wurmartige Körperform und die zahlreichen Stummelbeine. Eine Besonderheit vieler Lobopoden sind dagegen zahlreiche Panzerplatten, die Scleriten, die oft rückseitig den ganzen Körper mitsamt dem Kopf bedecken; da sie in langen, spitz zulaufenden Rückenstacheln auslaufen, hatten sie aller Wahrscheinlichkeit nach eine defensive Funktion zum Schutz vor Fressfeinden. Individuelle Scleriten lassen sich als Bestandteil der so genannten „small shelly fauna“ (SSF) schon vor Anbruch des Kambriums vor 543 Millionen Jahren nachweisen - demnach haben sich die Lobopoden beziehungsweise ihre unmittelbaren Vorfahren bereits in der Epoche des Ediacarium des Proterozoikums herausgebildet und waren somit die ersten auf Beinen laufenden Tiere.
Zu den bekannteren Gattungen zählt man etwa Aysheaia, die aus dem kanadischen Burgess-Schiefer bekannt ist und die von allen Lobopoden den modernen Stummelfüßern am ähnlichsten sieht - ein Paar Kopfanhänge lassen sich als Vorläufer der Antennen deuten. Xenusion war anscheinend in der Lage, sich mit den Stacheln nach außen zusammenzurollen, was Einblicke in die Verteidigungsstrategie der Lobopoden erlaubt. Die bei weitem berühmteste Lobopoden-Gattung ist hingegen Hallucigenia. Sie erhielt von dem Paläontologen Simon Conway Morris aufgrund ihres bizarren Aussehens ihren wissenschaftlichen Namen. Von ihm mit langen, stelzenförmigen Beinen und rätselhaften fleischigen Rückenauswüchsen rekonstruiert, galt sie lange als Paradebeispiel dafür, dass die Natur im Kambrium mit den verschiedensten und bizarrsten Körperbauplänen experimentierte. Es zeigte sich jedoch durch weitere Funde, dass die Rekonstruktion oben und unten vertauscht hatte; durch Interpretation der "Stelzen" als Rückenstacheln wurde klar, dass die fleischigen Auswüchse auf dem Rücken in Wirklichkeit Beine waren. Die zweite Rekonstruktion vertauschte jedoch ihrerseits Vorder- und Hinterseite des Tieres; erst nach weiteren Untersuchungen stellte sich heraus, dass Conway-Morris die Fossilien in dieser Hinsicht richtig orientiert hatte.
Lobopoden-Fossilien finden sich heute in Nordamerika, China und Nord- beziehungsweise Nordost-Europa. Die Fundorte lagen im Kambrium aufgrund der Kontinentaldrift jedoch wesentlich weiter südlich im tropischen und gemäßigten Gürtel der Erde und entsprachen damals flachen Lagunengewässern.
Die Lebensweise der Lobopoden liegt weitgehend im Dunkeln, einige Arten ernährten sich anscheinend carnivor, fraßen also andere Tiere wie etwa Schwämme (Porifera) oder seewalzenähnliche Stachelhäuter (Holothuroidea) der Gattung Eldonia. Dass sie sich wie moderne Stummelfüßer häuteten, gilt als nachgewiesen.
Stellten sie noch im Kambrium einen beträchtlichen Anteil an der Artenvielfalt, reduzierte sich diese drastisch im Ordovizium; spätestens mit dem Anbruch des Silurs gelten die Lobopoden im engeren Sinne als ausgestorben. Der US-amerikanische Paläontologe Stephen Jay Gould sieht dies als Symptom eines großflächigen Zusammenbruchs des Artenreichtums des Kambriums, die ein Ende der "Experimentierphase" ankündigte, seine Thesen gelten jedoch als umstritten.
Lobopoden werden traditionell aufgrund der sehr ähnlichen Morphologie mit den Stummelfüßern in Zusammenhang gebracht; nach dieser Ansicht waren sie Stammlinienvertreter dieses Taxons:
Stummelfüßer (Onychophora, alternativ: Lobopodia) |==diverse Lobopoden-Gattungen (meereslebend, paraphyletisch) |--Udeonychophora (alternativ: Onychophora) (landlebend, umfassen moderne und fossile Arten)
Gelegentlich bezeichnet der wissenschaftliche Name Onychophora nur die landlebenden Formen, während die größere Gruppe, die auch die verschiedenen Lobopoden einschließt dann als Lobopodia bezeichnet wird.
Eine wichtige Alternativsicht geht davon aus, dass die Lobopoden stattdessen Stammlinienvertreter der Panarthropoda und Tactopoda waren:
Panarthropoda |--Aysheaia |--N. N. |--Stummelfüßer (Onychophora) inklusive ausgestorbener landlebender Arten |--N. N. |==diverse Lobopoden-Gattungen (paraphyletisch) |--Tactopoda (Bärtierchen + Gliederfüßer)Die Gattung Aysheaia, die in besonders vielen Merkmalen mit den modernen Stummelfüßern übereinstimmt, bildet demnach die Schwestergruppe aller anderen Panarthropoda, die sich sodann in die Stummelfüßer und alle restlichen Formen aufteilen. Die verbliebenen Lobopoden-Gattungen gelten nun als Vertreter derjenigen Entwicklungslinie, die zu den modernen Bärtierchen und Gliederfüßern hinführt. Aus der kladistischen Analyse lässt sich dann ein Trend hin zu verstärkter äußerer Segmentierung und zu einer Abstimmung der feinen Körper- und Beinringung auf die Lage der Segmentgrenzen feststellen.
Das Resultat lässt sich allerdings nur schwer mit der Articulata-Hypothese vereinbaren, weil es bedeutet, dass die ausgeprägte Segmentierung von Ringelwürmern und Gliederfüßern unabhängig voneinander entstanden sein muss. Für die Stummelfüßer hieße dies, dass das Fehlen einer deutlich erkennbaren äußerlichen Segmentierung ein ursprüngliches, „primitives“ und kein sekundäres Merkmal darstellt. Die zweite Hypothese zur systematischen Stellung der Lobopoden verträgt sich daher weitaus besser mit dem oben angesprochenen Ecdysozoa-Konzept.
Landlebende Formen
Vier terrestrische Formen der Stummelfüßer sind bekannt, bei denen es im Gegensatz zu den Lobopoden über die systematische Stellung keinen großen Zweifel gibt.
Wann sich der Übergang an Land vollzog, ist unbekannt; als plausibel gilt, dass er zwischen Ordovizium und spätem Silur, also vor etwa 490 bis 420 Millionen Jahren, in der Gezeitenzone stattfand. Die typische Panzerung der Lobopoden könnte, falls sie bei den Vorfahren der modernen Arten jemals vorhanden war, zu diesem Zeitpunkt verloren gegangen sein, vielleicht um sich durch die gewonnene Flexibilität besser in engen Lebensräumen bewegen zu können. In der Gezeitenzone könnten sich auch die verschiedenen Anpassungen an das Landleben herausgebildet haben, etwa die Tracheen, die demnach unabhängig von denen der Insekten entstanden wären oder der interne Befruchtungsmodus, der auch bei Wassermangel eine Übertragung der Samenzellen in eine lebensfreundliche Umgebung möglich macht. Über den Zeitpunkt der Umfunktionierung der Nephridien in Speichel- und genitale Hilfsdrüsen lässt sich nur spekulieren; bei den zum Beutefang eingesetzten Schleimdrüsen ist immerhin bekannt, dass sie unter Wasser funktionsunfähig sind, woraus vorsichtig geschlossen werden kann, dass sie sich wohl erst an Land ausbildeten, vielleicht ursprünglich zur Abwehr von Fressfeinden.
Wo immer der Übergang letztlich stattfand, es war anscheinend keine die Fossilienbildung begünstigende Umgebung; jedenfalls existiert aus dieser Frühzeit kein einziges Fossil. Die aus dem tropischen Gürtel des späten Karbon bekannte Art Helenodora inopinata war sehr wahrscheinlich bereits landlebend und unterscheidet sich nur wenig von den modernen Formen. Aus diesem Grund und der immer noch großen Ähnlichkeit zu Lobopoden wie Aysheaia gelten Stummelfüßer als Paradebeispiel für evolutionäre Stasis und eine bradytelische Evolutionsrate, bei der sich die biologische Organisation des Gesamtorganismus nur sehr langsam wandelt, weil starke stabilisierende Selektion die gesamte Entwicklungslinie in einem engen Korridor des morphologisch-anatomischen „Raums“ eingrenzt und größere Abweichungen von der „typischen“ Stummelfüßerform nicht toleriert.
Aus dem Erdmittelalter ist nur ein Fossil bekannt, die kreidezeitliche Art Cretoperipatus burmiticus, die in Bernstein aus dem südostasiatischen Myanmar gefunden wurde und auf ein Alter von 100 Millionen Jahren datiert wird. Sie kann sogar bereits einer der modernen Familien, den Peripatidae, zugeordnet werden.
Da heutige Stummelfüßer eine so genannte Gondwana-Verteilung aufweisen, spricht vieles dafür, dass ihr letzter gemeinsamer Vorfahre auf eben diesem Urkontinent lebte. Dass auch außerhalb Gondwanas oder der heutigen Überreste dieses Kontinents landlebende Stummelfüßer existierten, beweist die Existenz von Succinipatopsis balticus, einer 44 Millionen Jahre alten Art, die sich in baltischem Bernstein aus der Periode des unteren Eozäns erhalten hat. Das Verbreitungsgebiet der Stummelfüßer muss also einstmals wesentlich größer gewesen sein als heute - wann und aus welchem Grund alle nicht-Gondwana-Arten ausstarben, ist unbekannt.
Die vierte fossile Art ist Tertiapatus dominicanus, die aus Bernstein von der Karibik-Insel Hispaniola bekannt ist. Es ist möglich, aber nicht beweisbar, dass sie schon einer der beiden modernen Familien, den Peripatidae, angehörte. Die Art lässt sich auf ein Alter von 17 bis 20 Millionen Jahren datieren.
An beiden Fossilien des Tertiärs lassen sich deutlich die Schleimdrüsen erkennen, die sich somit spätestens zu diesem Zeitpunkt, wahrscheinlich aber schon wesentlich früher herausgebildet haben müssen. Formell werden die beiden tertiären Formen in einem Taxon Tertiapatoidea zusammengefasst, das aber vermutlich nur ein Formtaxon darstellt.
Systematik
Die modernen Stummelfüßer bilden eine so genannte monophyletische Gruppe, umfassen also alle Nachkommen ihres gemeinsamen Vorfahrens. Wichtige gemeinsame abgeleitete Merkmale (Synapomorphien) sind zum Beispiel die Kieferwerkzeuge (Mandibeln) des zweiten und die Mundpapillen mit zugehörigen Schleimdrüsen des dritten Körpersegments, bauchseitig verlaufende Nervenstränge mit zahlreichen Querverbindungen pro Segment und die spezielle Form der Tracheen.
Bis 2004 wurden etwa 155 moderne Arten beschrieben, die 47 Gattungen zugeordnet werden; die tatsächliche Artenzahl ist aber wahrscheinlich etwa doppelt so groß. Am bekanntesten ist die Typgattung Peripatus, die bereits 1825 beschrieben wurde und im englischsprachigen Raum auch stellvertretend für alle Stummelfüßer steht.
Alle Gattungen werden einer von zwei Familien zugeordnet, deren Verbreitungsgebiete nicht überlappen, sondern durch aride Gebiete oder Meere voneinander getrennt sind.
- Die Peripatopsidae weisen verhältnismäßig viele als ursprünglich oder "primitiv" eingeschätzte Merkmale auf. Sie besitzen 13 bis 25 Beinpaare; hinter oder zwischen dem letzten davon befindet sich die Geschlechtsöffnung (Gonopore). Sowohl eierlegende (ovipare), eierlebendgebärende (ovovivipare) als auch echt lebendgebärende (vivipare) Arten kommen vor, eine Plazenta besitzen Peripatopsidae allerdings grundsätzlich nicht. Ihr Verbreitungsgebiet liegt zirkumaustral in Australien und Tasmanien, Neuseeland, Neu-Guinea, Südafrika und Chile.
- Die Peripatidae zeigen eine Reihe von abgeleiteten Merkmalen. Von größerer Durchschnittslänge als die Peripatopsidae besitzen sie auch mehr Beinpaare: Deren Zahl variiert innerhalb der Gruppe zwischen 22 und 43; die Gonopore liegt immer zwischen dem vorletzten davon. Ovipare Arten gibt es bei den Peripatidae nicht, die weitaus überwiegende Anzahl ist vivipar. Bei vielen Weibchen der letzteren Gruppe bildet sich eine Plazenta aus, die den heranwachsenden Embryo mit Nährstoffen versorgt. Vorkommen der Peripatidae sind auf die tropische und subtropische Zone beschränkt; im Einzelnen kommen sie in Mittel- und im nördlichen Südamerika sowie auf mehreren karibischen Inseln vor, daneben auch in Westafrika, Nordindien, Malaysia und einzelnen indonesischen Inseln.
Quellen
Literatur
- H. Ruhberg: Onychophora, Stummelfüßer. in: W. Westheide, R. Rieger : Spezielle Zoologie. Teil 1. Einzeller und Wirbellose Tiere. Ulmer, Stuttgart 1974, Elsevier, München 2007, ISBN 3800124297, ISBN 3-8274-1575-6
- Edward E. Ruppert, R. S. Fox, R. D. Barnes: Invertebrate Zoology: A functional evolutionary approach. Kapitel 15. Brooks/Cole, London 2004, S.505, ISBN 0-03-025982-7
- R. C. Brusca, G. J. Brusca: Invertebrates. Kapitel 15. Sinauer Associates, Sunderland Mass 2003, S. 463, ISBN 0878930973
Wissenschaftliche Literatur
- G. E. Budd: The morphology of Opabinia regalis and the reconstruction of the arthropod stem group in: Lethaia. Taylor & Francis, Oslo 29.1996, S. 1. ISSN 0024-1164
- J. Eriksson: Evolution and Development of the Onychophoran Head and Nervous System. in: Comprehensive summaries of Uppsala dissertations from the Faculty of Science and Technology. Upsala 833, 2003. ISSN 1104-232X
- X. Hou, J. Bergström: Cambrian lobopodians - ancestors of extant onychophorans. in: Zoological Journal of the Linnean Society. Blackwell, Oxford 114.1995, S. 3. ISSN 0024-4082
- J. Monge-Najera: Phylogeny, biogeography and reproductive trends in the onychophora. in: Zoological Journal of the Linnean Society. Blackwell, Oxford 114.1995, S. 21. ISSN 0024-4082
- T. R. New: Onychophora in invertebrate conservation: priorities, practice and prospects. in: Zoological Journal of the Linnean Society. Blackwell, Oxford 114.1995, S. 77. ISSN 0024-4082
- G. Poinar: Fossil onychophorans from Dominican and Baltic Amber: Tertiapatus dominicanus n.g., n.sp. (Tertiapatidae n. fam.) and Succinipatopsis balticus n.g., n. sp. (Succinipatopsidae n. fam.) with a proposed classification of the subphylum Onychophora. in: Invertebrate Biology. Blackwell, Malden Mass 119.2000, S. 104. ISSN 1077-8306
- G. Poinar: Fossil Velvet Worms in Baltic and Dominican Amber: Onchychophoran Evolution and Biogeography. in: Science. Moses King, Cambridge Mass 273.1996, S. 1370. ISSN 0096-3771
- C. J. Wedeen, R. G. Kostriken, D. Leach, P. Whitington: Segmentally iterated expression of an engrailed-class gene in the embryo of an Australian onychophoran. in: Development Genes and Evolution. Springer, Heidelberg 207.1997,4, S. 282. ISSN 0949-944X
Weblinks
- Galerie (Englisch)


Wikimedia Foundation.