- Humanes Immundefizienz-Virus
-
Humanes Immundefizienz-Virus 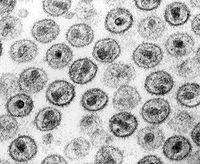 HI-Viren im TEM
HI-Viren im TEM
Systematik Reich: Viren Unterfamilien: Orthoretrovirinae Gattung: Lentivirus Art: Humanes Immundefizienz-Virus Taxonomische Merkmale Genom: (+)ssRNA linear, dimer Symmetrie: komplex Hülle: vorhanden Wissenschaftlicher Name Human immunodeficiency virus (engl.) Taxon-Kurzbezeichnung HIV-1, HIV-2 Links NCBI Taxonomie: 11676 NCBI Reference: AF033819 ICTVdB Virus Code: 00.061.1.06.009 Das Humane Immundefizienz-Virus (englisch human immunodeficiency virus), zumeist abgekürzt als HIV oder auch bezeichnet als Menschliches Immunschwäche-Virus oder Menschliches Immundefekt-Virus, ist ein Virus, das zur Familie der Retroviren und zur Gattung der Lentiviren gehört. Eine Ansteckung führt nach einer unterschiedlich langen, meist mehrjährigen Inkubationszeit zu AIDS (engl. acquired immunodeficiency syndrome ‚erworbenes Immundefizienzsyndrom‘), einer derzeit noch unheilbaren Immunschwächekrankheit.
Die Verbreitung von HIV hat sich in den letzten 25 Jahren zu einer Pandemie entwickelt, die nach Schätzungen der Organisation UNAIDS bisher etwa 25 Millionen Leben gefordert hat. Etwa 33,4 Millionen Menschen sind weltweit mit dem Virus infiziert, davon ca. 50 Prozent Männer. Die Zahl der Neuinfektionen sinkt seit 1996 stetig und lag 2008 bei 2,7 Millionen Menschen.[1] In Deutschland leben etwa 70.000 Menschen mit HIV, davon 81 Prozent Männer und 200 Kinder. Im Jahr 2009 war AIDS bei 11.300 Personen bereits ausgebrochen; außerdem kam es zu 3000 neuen HIV-Infektionen.[2] In Österreich infizierten sich 2005 insgesamt 453 Menschen mit HIV. In der Schweiz leben etwa 20.000 HIV-Infizierte und etwa 2700 AIDS-Kranke.[3]
Inhaltsverzeichnis
Struktur und Aufbau des HI-Virus
 Aufbau des HI-Virions
Aufbau des HI-VirionsHIV gehört zu den komplexen Retroviren, das heißt, sie besitzen neben den kanonischen retroviralen Genen gag, pol und env weitere regulatorische und akzessorische Leseraster, namentlich bei HIV-1 tat, rev, vif, vpu, vpr und nef.
Das Viruspartikel hat einen Durchmesser von etwa 100 bis 120 Nanometer und ist von einer Lipoproteinhülle umgeben. Eingebettet in diese Hülle sind pro Virion etwa 10 bis 15 (14±7) sogenannte Spikes (dt. Dornen), ca. 10 Nanometer große env-Glykoproteinkomplexe; die Dichte der Spikes ist somit recht niedrig, hätten doch 73±25 dieser Fortsätze auf der Oberfläche eines HIV-Partikels Platz. Ein HIV-Spike besteht aus zwei Untereinheiten: Je drei Moleküle des externen Oberflächen-(engl. surface)-Glykoproteins gp120 sind nicht-kovalent an drei Moleküle des transmembranen Hüll-(engl. envelope)-Glykoproteins gp41 gebunden.[4][5][6][7] Gp120 ist für die Bindung des Virus an die CD4-Rezeptoren der Zielzellen von entscheidender Bedeutung. Da die Hülle des HI-Virus aus der Membran der Wirtszelle entsteht, befinden sich in ihr ebenfalls verschiedene Proteine der Wirtszelle, zum Beispiel HLA Klasse I und II Moleküle sowie Adhäsionsproteine.
Mit der Innenseite der Membran sind die durch gag kodierten Matrixproteine assoziiert. Im Inneren des Virions findet sich das Viruskapsid (oder core), das aus den durch gag kodierten Kapsidproteinen aufgebaut ist. Im Kapsid findet sich, an die durch gag kodierten Nukleokapsidproteine assoziiert, das virale Genom (9,2 kb) in Form zweier Kopien der einzelsträngigen RNA in Plusstrangorientierung. Ebenso befinden sich im Kapsid die Enzyme reverse Transkriptase (RT), Integrase sowie einige der akzessorischen Proteine. Die Protease ist maßgeblich beteiligt an der Partikelbildung und findet sich daher im gesamten Partikel.
Geschichte
HIV ist die vom International Committee on Taxonomy of Viruses 1986 empfohlene Bezeichnung. Es ersetzt die ehemaligen Benennungen wie Lymphadenopathie-assoziiertes Virus (LAV), humanes T-Zell-Leukämie-Virus III (HTLV-III) oder AIDS-assoziiertes Retrovirus (ARV).
HIV Typ 1 wurde 1983 zum ersten Mal von Luc Montagnier und Françoise Barré-Sinoussi vom Institut Pasteur in Paris beschrieben.[8] In derselben Ausgabe des Journals Science veröffentlichte Robert Gallo, der Leiter des Tumorvirus-Labors am NIH, ebenfalls die Entdeckung eines Virus, das seiner Meinung nach AIDS auslösen könnte.[9] Er beschreibt in dieser Veröffentlichung jedoch die Isolierung von Humanen T-Zell-Leukämie-Viren Typ I (HTLV-1; kurze Zeit später auch von HTLV-2) bei AIDS-Patienten, die in seinen Proben zufällig neben dem HI-Virus vorlagen, und isolierte erst etwa ein Jahr später auch das HI-Virus. Dennoch beanspruchten sowohl Gallo und Montagnier jeweils die Erstentdeckung für sich. Daraufhin folgte ein jahrelanger Rechtsstreit, bei dem es auch um das Patent für den neu entwickelten HIV-Test ging. 1986 wurde HIV-2 entdeckt.
Die beiden Forscher Françoise Barré-Sinoussi und Luc Montagnier wurden 2008 für die Entdeckung des HI-Virus mit dem Nobelpreis für Medizin ausgezeichnet.[10]
Im Mai 2005 gelang einem internationalen Forscherteam erstmals der Nachweis, dass der Ursprung von HIV beim Affen liegt. Das Forscherteam nahm dazu in der Wildnis des zentralafrikanischen Kamerun 446 Kotproben freilebender Schimpansen. Etliche Proben wiesen Antikörper gegen Simianes Immundefizienz-Virus (kurz SIV; engl. Simian Immunodeficiency Virus) auf, die Schimpansenversion des HI-Virus, wie das Team im US-Fachjournal Science veröffentlichte. Zwölf Proben waren fast identisch mit dem HIV-1 bei Menschen. Das Team betonte, dass die Antikörper zuvor nur bei Schimpansen in Gefangenschaft nachgewiesen wurden. Ursprüngliche Quelle des HI-Virus sind die Schimpansen jedoch nicht. Sie sollen sich im westlichen Zentralafrika mit SIV oder einem Vorläufer dieses Virus’ bei anderen Affenarten infiziert haben. Etwa im 20. Jahrhundert infizierten sich erstmals Menschen mit dem SIV, das anschließend in deren Organismen zum AIDS verursachenden HIV mutierte. Damit hat der AIDS-Erreger bereits mindestens zweimal die Artengrenze übersprungen, nämlich vom Affen zum Menschenaffen und anschließend zum Menschen. Wie das Virus auf den Menschen übertragen wurde, ist unklar. Man geht davon aus, dass Jäger, die Affen gejagt und verspeist haben, mit dem Virus erstmals infiziert wurden.
Eine andere These war, dass ein Impfstoff gegen Poliomyelitis (Kinderlähmung) im Jahre 1959 durch Affen, die das Virus trugen, verunreinigt worden sei. Nach dieser These wurden im ehemaligen Belgisch-Kongo Schimpansennieren zur Vermehrung des Impfstoffes verwendet und anschließend Hunderttausende Menschen durch eine Schluckimpfung geimpft, wodurch SIV auf den Menschen übertragen worden und zum HIV mutiert sei.[11][12][13] Allerdings zeigte eine Analyse der Mutationen, dass mit 95-prozentiger Wahrscheinlichkeit der Ursprung des Stammes HIV-1 vor das Jahr 1930 zu datieren ist.[14] Im Februar 2000 wurde eine Probe der verteilten Schluckimpfungen gefunden und untersucht. In dieser zeigten sich weder Spuren von HIV noch von SIV.[15]
Der älteste anhand von Blutproben gesicherte Nachweis einer HIV-Infektion stammt aus Zaire und aus dem Jahre 1959.[16] Um 1966 soll das Virus nach Haiti und von dort aus 1969 in die USA gelangt sein.
Neuere Untersuchungen kommen zu dem Ergebnis, dass das HI-Virus schon um 1908 zum ersten Mal einen Menschen infiziert hat, wobei aufgrund neuester statistischer Analysen (sogenannte molekulare Uhr) das Zeitfenster für das Erstauftreten mit hoher Wahrscheinlichkeit auf die Jahre zwischen 1902 und 1921 eingegrenzt werden kann.[17][18][19][20][21]
Einteilung und Systematik
Es sind bisher zwei verschiedene Arten von HI-Viren bekannt, die als HIV-1 und HIV-2 bezeichnet werden. Die Homologie zwischen HIV-1 und HIV-2 beträgt auf Aminosäuresequenzebene nur etwa 45 bis 50 Prozent. Sie können weiter in Subtypen unterteilt werden, die teilweise mit unterschiedlicher Häufigkeit in verschiedenen Regionen der Welt auftreten. In Mitteleuropa ist zum Beispiel der Subtyp B aus der Gruppe M von HIV-1 am häufigsten, besonders unter Homosexuellen und Drogenabhängigen. HIV-1, das insgesamt häufiger ist, und HIV-2 ähneln sich prinzipiell hinsichtlich des klinischen Infektionsverlaufs und der krankmachenden (pathogenen) Eigenschaften, auch wenn die Infektion mit HIV-2 wohl insgesamt langsamer verläuft. Die beiden Stämme sehen unter dem Elektronenmikroskop gleich aus, unterscheiden sich jedoch in der molaren Masse der Proteine und in der Anordnung und Nukleotidsequenz der Gene. HIV-1 und HIV-2 entstanden aus unterschiedlichen Typen der bei bestimmten Affenarten vorkommenden SI-Viren.
Von HIV-1 gibt es vier Untergruppen, die mit M, N, O und P bezeichnet werden.[22][23] M steht für major group (engl. für Hauptgruppe) und ist am häufigsten, die O-Gruppe wurde nach outlier (Sonderfall) benannt und das N der Gruppe N steht für new (neu). In die Gruppe M von HIV-1 fallen mehr als 90 Prozent aller HIV-Infektionen und ist verantwortlich für die Infektion von weltweit bisher rund 60 Millionen Menschen (Stand 2010).[24] Diese Gruppe wird wiederum in neun Subtypen unterteilt, die mit A, B, C, D, F, G, H, J und K bezeichnet werden. Die häufigsten sind die Subtypen B (kommt vor allem in Nordamerika und Europa vor), A und D (vor allem in Afrika) und C (hauptsächlich in Afrika und Asien). Eine Koinfektion mit verschiedenen Subtypen kann dazu führen, dass rekombinante Formen entstehen, die circulating recombinant forms (CRFs) genannt werden. Die Klassifikation der HIV-Stämme ist entsprechend komplex und noch nicht abgeschlossen.
Die HIV-1-Gruppe O scheint bisher fast ausschließlich in Westafrika verbreitet zu sein, während die neu entdeckten Gruppen N- und P-Viren bislang nur bei einigen wenigen Menschen nachgewiesen werden konnten.[24]
Heute ist bekannt, dass der Haupterreger von AIDS, HIV-1, ursprünglich von SIV-infizierten Schimpansen und Gorillas auf den Menschen übertragen wurden. So konnte im Falle von HIV-1 für die Gruppen M und N nachgewiesen werden, dass sie aus Schimpansen stammen, während HIV-1 P von Gorillas auf den Menschen übertragen wurde.[22] Ob HIV-1 O ursprünglich aus SIV-infizierten Schimpansen oder Gorillas stammt ist noch nicht abschließend geklärt. Alle vier HIV-1-Gruppen (M, N, O und P) sind auf vier unabhängige Zoonosen (d. h. Tier-zu-Mensch-Übertragungen) zurückzuführen, wobei bislang nur die Gruppe M die Ausmaße einer Pandemie angenommen hat.[25][24] Der Grund für die hohe Infektiösität des HIV-1 M-Stammes beruht unter anderem auf spezifischen Eigenschaften des Virusproteins Vpu der M-Gruppe, mit deren Hilfe gleich zwei Infektionsbarrieren überwunden werden: zum einen wird der antivirale Faktor Tetherin auf humanen Zellen effektiv ausgeschaltet und zum anderen wird der CD4-Rezeptor abgebaut.[26][24][27]
Epidemiologie
Die HIV-Prävalenz bei Erwachsenen (15 bis 49 Jahre) für Zentral- und Westeuropa lag 2008 bei 0,3 Prozent, die weltweite HIV-Prävalenz bei 0,8 Prozent. Im subsaharischen Afrika (5,2 Prozent) und in der Karibik (1,0 Prozent) war sie überdurchschnittlich hoch. In einzelnen Staaten, wie zum Beispiel Swasiland, Botsuana oder Lesotho sind ca. ein Viertel der 15- bis 49-Jährigen mit dem HI-Virus infiziert. Stark unterdurchschnittlich war die HIV-Prävalenz im Jahr 2008 in den Regionen Ostasien (< 0,1 Prozent) sowie in Nordafrika und dem Mittleren Osten (0,2 Prozent). Die Zahl der Neuinfektionen lag 2008 bei ca. 30.000 Betroffenen in Zentral- und Westeuropa. [28]
Übertragung
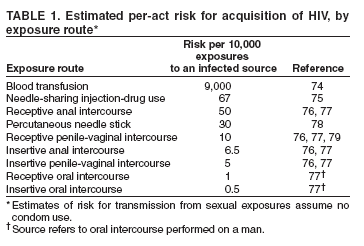
Geschätztes Übertragungsrisiko pro Akt, je nach Infektionsweg (bei sexuellen Kontakten ohne Kondom)[29] Infektionsweg Risiko pro 10000 Kontakten mit infektiöser Quelle In Prozent Bluttransfusion 9000 90 % Nadel teilen bei Drogeninjektion 67 0,67 % Empfangender Analverkehr 50 0,50 % Nadelstich durch die Haut 30 0,30 % Empfangende vaginale Penetration mit Penis 10 0,10 % Einführender Analverkehr 6,5 0,065 % Einführender Vaginalverkehr 5 0,05 % Empfangender Oralverkehr 1 0,01 % Einführender Oralverkehr 0,5 0,005 % Das HI-Virus wird durch Kontakt mit den Körperflüssigkeiten Blut, Sperma (auch Präejakulat), Vaginalsekret sowie Muttermilch und Liquor cerebrospinalis übertragen. Potenzielle Eintrittspforten sind frische, noch blutende Wunden und Schleimhäute (Bindehaut, Vaginal-[30] und Analschleimhaut) bzw. nicht ausreichend verhornte, leicht verletzliche Stellen der Außenhaut (Eichel, Innenseite der Vorhaut, Anus). Als häufigste Infektionswege sind zu nennen der Anal- oder Vaginalverkehr ohne Verwendung von Kondomen und die Benutzung unsteriler Spritzen beim intravenösen Drogenkonsum. Oralverkehr gilt nach jüngsten Studien als weniger infektiös. Eine Ansteckung ist bei Oralverkehr vor allem möglich, wenn Sperma oder Menstruationsblut auf die Mundschleimhaut gelangt. Homosexuelle Männer gelten als Risikogruppe, da Analverkehr bei ihnen stärker verbreitet ist als in der Gruppe der Heterosexuellen. Wie hoch das Risiko beim Geschlechtsverkehr ist, hängt vor allem von der Viruskonzentration in der Samenflüssigkeit und im Scheidensekret sowie von der Viruslast im Blut ab. Diese ist unmittelbar nach der Infektion, bevor sich ausreichend Antikörper gebildet haben, besonders hoch, nimmt dann aber zunächst ab und steigt in späten Stadien der Erkrankung wieder an.[31] Wie sich bei aktuellen Studien gezeigt hat, ist das Infektionsrisiko für beschnittene Männer etwas geringer. Die Beschneidung hinterlässt, nach der gängigsten Annahme, durch die Entfernung der Vorhaut eine geringere Angriffsfläche für das Virus.[32] Aus diesem Grund empfahl die Weltgesundheitsorganisation (WHO) 2007 ihren Mitgliedsländern die Beschneidung als Präventivmaßnahme zur Eindämmung von AIDS, wofür sie allerdings von Experten kritisiert wurde.
Bluttransfusionen sind ebenfalls eine mögliche Infektionsquelle: das Risiko für den Empfänger sich durch mit HIV kontaminiertes Blut zu infizieren wird auf 90% geschätzt.[33] So kam es Anfang der 1980er Jahre in vielen Ländern zu verschiedenen Blutskandalen. Diese Ansteckungsmöglichkeit hat heute in Deutschland wegen der 1985 eingeführten Routine-Untersuchungen auf HIV-Antikörper der Blutspender allerdings kaum noch Bedeutung.[34] Da zwischen der Ansteckung des Spenders und der Nachweisbarkeit von Antikörpern im HIV-Test in Einzelfällen bis zu drei Monate verstreichen können (diagnostische Lücke), werden seit Anfang 2002 zwingend alle deutschen Blutspenden auch mittels Polymerase-Kettenreaktion (PCR) auf die Anwesenheit des Virus getestet.
Das Risiko der Infektion eines Kindes durch eine HIV-infizierte Mutter während der Schwangerschaft oder während der Geburt wird ohne Behandlung auf 15 bis 30 Prozent geschätzt. Eine Übertragung des Virus beim Stillen ist ebenfalls möglich. Bei bekannter HIV-Infektion der Mutter kann das Risiko einer Übertragung auf das Kind durch die Gabe antiretroviraler Medikamente, durch eine Geburt mittels Kaiserschnitt und durch Abstillen auf ein bis zwei Prozent vermindert werden.[35]
Die sogenannte CHAT-Survey-Studie[36] des schweizerischen Bundesamtes für Gesundheitswesen (BAG) – eine Nachbefragung von Menschen, die im Verlauf eines Jahres positive HIV-Tests erhielten – ergab, dass 49 Prozent aller Neuinfizierten die Infektion von ihrem festen Sexualpartner erhielten; 38 Prozent wurden von einem zwar bekannten, aber nicht festen Gelegenheitspartner infiziert. 10 Prozent der neuinfizierten Personen wussten schon vorher, dass ihr Partner HIV-positiv ist. Wurde jemand von seinem bereits infizierten Partner absichtlich angesteckt, spricht man vom sogenannten Pozzen. Nur 13 Prozent der Heterosexuellen steckten sich bei anonymen sexuellen Begegnungen an. Bei Homosexuellen spielten die Infektionen durch feste Partner eine geringere Rolle – anonyme Sexualkontakte machten 26 Prozent der Infektionen aus.[37]
Eine Möglichkeit, sich durch Zungenküsse anzustecken, besteht nur dann, wenn blutende Wunden, beispielsweise Verletzungen des Zahnfleisches, im Mund vorhanden sind. Die HIV-Konzentration in Tränen, Schweiß und Speichel reicht für eine Ansteckung nach heutigem Erkenntnisstand nicht aus. Außerdem lässt die AIDS-Epidemiologie eine Infektion durch Insektenstiche oder durch Tröpfcheninfektion äußerst unwahrscheinlich erscheinen.
Menschen, die einer akuten Ansteckungsgefahr ausgesetzt waren, sollten möglichst bald (idealerweise innerhalb von zwei Stunden) einen Arzt aufsuchen, um sich beraten zu lassen und gegebenenfalls eine Postexpositionelle Prophylaxe (PEP) durchzuführen. Nach Ablauf von 48 bzw. 72 Stunden wird eine medikamentöse PEP nicht mehr als sinnvoll erachtet.
Hinsichtlich der Infektionswahrscheinlichkeiten siehe ausführlich unter AIDS.
Vermehrungszyklus des HIV
Zur Vermehrung benötigt das Virus Wirtszellen, die den CD4-Rezeptor auf der Oberfläche tragen. Dies sind vor allem die CD4-tragenden T-Lymphozyten (T-Helferzellen), die beim Menschen eine wichtige Rolle sowohl in der zellulären als auch in der humoralen Immunabwehr spielen und unter anderem die Antikörperbildung unterstützen. Neben T-Lymphozyten besitzen auch Monozyten, Makrophagen und dendritische Zellen CD4-Rezeptoren.
Fusion mit der Wirtszelle
Um mit der Zellmembran der Wirtszelle verschmelzen zu können, binden die Oberflächenproteine gp120 an die CD4-Rezeptoren. Durch die Bindung kommt es zu einer Konformationsänderung im Transmembranprotein gp41, ein Mechanismus, der einer „Schnappfeder“ oder einer „Mausefalle“ ähnelt. Der Wirkstoff Enfuvirtid bzw. T20 ist ein Peptid, das die Konformationsänderung blockiert und somit die Anheftung des Virus erschwert (siehe unten).
Neben den CD4-Rezeptoren sind weitere Co-Rezeptoren an der Bindung des HI-Virus an weißen Blutzellen beteiligt:[38] Die Chemokin-Rezeptoren CCR5 an monozytären Zellen und CXCR4 an T-Zellen sind an der Bindung beteiligt.[39][40] Die unterschiedliche Ausprägung dieser Rezeptoren beeinflusst die Ansteckungswahrscheinlichkeit und den Verlauf der HIV-Infektion.[41] Moleküle, die die CCR5 Rezeptoren blockieren sollen, wurden getestet.[42][43] Derzeit ist der CCR5 Rezeptorblocker Maraviroc seit September 2007 zugelassen.[44] Die Zulassung ist jedoch unter folgenden Einschränkungen erfolgt: „Einsatz nur beim Nachweis von Viren, die ausschließlich über den CCR5-Rezeptor in die Zelle eintreten; vor Einsatz des Medikaments ist daher ein Test (Blutabnahme) erforderlich. Nicht für die Ersttherapie zugelassen.“
Ebenso sind Menschen, die homozygot die sogenannte Delta-32-Mutation des CCR5-Corezeptor-Gens aufweisen, schwerer mit HIV infizierbar. Dies trifft auf etwa ein Prozent der Bevölkerung in Europa zu. In geringerem Maße trifft dies auch auf Mutationen des CCR2-Gens zu. Menschen mit HLA B27/B57 (siehe Human Leukocyte Antigen) zeigen einen langsameren Verlauf der Erkrankung.
Einbau des HI-Virus-Genoms in die Wirtszelle
 Vermehrung des HIV
Vermehrung des HIVDas HIV baut zur Vermehrung seine Erbsubstanz, die bei ihm in Form eines RNA-Genoms vorliegt, nach der so genannten reversen Transkription in die doppelsträngige DNA des Genoms der Wirtszelle ein. Die Umwandlung von viraler RNA in provirale DNA im Cytoplasma der Wirtszelle durch das Enzym Reverse Transkriptase ist ein entscheidender Schritt im Reproduktionszyklus der Retroviren. Da es kein menschliches Gen gibt, das für eine Reverse Transkriptase kodiert und dieses Enzym nur bei Viren wie Retro- und Hepadnaviren vorkommt, stellt die Reverse Transkriptase ein wichtiges Ziel therapeutischer Intervention dar und ist Ansatzpunkt zweier pharmakologischer Wirkstoffklassen.
Nach reverser Transkription und Transport in den Zellkern schließt sich die Integration des Virus-Genoms in das menschliche Erbgut durch ein weiteres virales Enzym, die Integrase, an. In neueren Arbeiten wurde gezeigt, dass die virale DNA schon vor der Integration abgelesen wird und virale Proteine gebildet werden. Demnach liegt die HIV-DNA als integrierte und nicht-integrierte Form vor. Auch existieren zirkuläre Formen von HIV-DNA.
Das nun als integriertes Provirus vorliegende virale Genom zeigt einen charakteristischen Aufbau, wobei die kodierenden Bereiche auf beiden Seiten von identischen regulatorischen Sequenzen, die im Verlauf der reversen Transkription generiert wurden, den sogenannten LTRs, flankiert sind. Der Promotor, unter dessen Kontrolle die Transkription der verschiedenen mRNAs erfolgt, liegt im Bereich des LTR und wird durch das virale Protein Tat aktiviert. Eine ungespleißte RNA dient als virales Genom für die nächste Generation von HI-Viren sowie als mRNA für die Translation eines gag- sowie mittels einer in einem von 20 Fällen vorkommenden Verschiebung des Leserasters eines gag-pro-pol-Vorläuferproteins. Gespleißte RNAs kodieren für env sowie die ebenfalls im 3'-Bereich befindlichen weiteren Proteine.
Im weiteren Verlauf folgt die Morphogenese, das heißt, über verschiedene Interaktionen finden die viralen Bestandteile wie gag-, pro-pol- und env-Vorläuferproteine sowie die RNA zusammen und formen sich zu zunächst unreifen Virionen, die sich von der Plasmamembran abschnüren. Durch weitere Reifungsprozesse entsteht das reife Viruspartikel, bereit für die Infektion der nächsten Zelle. Zu den Reifungsprozessen gehört insbesondere die Spaltung der Vorläuferproteine, teils durch die virale Protease, teils durch zelluläre Enzyme, in ihre einzelnen Bestandteile, also von Gag in Matrix-, Kapsid- und Nukleokapsidprotein, Pol in Protease, Reverse Transkriptase mit RNase H und Integrase sowie Env in Oberflächen- und Transmembraneinheit.
Das Virus in infizierten und ruhenden CD4-positiven T-Zellen entzieht sich dem Angriff seitens antiviraler Medikamente und des Immunsystems. Zu einer Aktivierung dieser Immunzellen kommt es nach Antigenkontakt, zum Beispiel im Rahmen gewöhnlicher oder einer opportunistischen Infektion. Während die Zelle eigentlich gegen einen anderen Krankheitserreger vorgehen will, beginnt sie statt dessen Virusproteine zu produzieren und neue Viren freizusetzen. Diese infizieren dann wiederum andere Zellen.
Was das HI-Virus so außergewöhnlich überlebensfähig macht, ist seine Wandlungsfähigkeit oder, besser gesagt, seine schnelle Evolutionsrate. Von den Influenza-Viren (Grippe) zum Beispiel entwickeln sich in derselben Zeit auf der ganzen Welt nicht einmal halb so viele neue Unterarten wie vom HI-Virus in einem einzelnen infizierten Menschen.
Die lange Inkubationszeit von zehn Jahren ist ein Problem, da viele Infizierte unter Umständen noch jahrelang andere Personen infizieren, bevor ihre Infektion erkannt oder von ihnen selbst bemerkt wird.
Verlauf der HIV-Infektion
 HI-Viren sammeln sich vor dem Verlassen der Immunzelle an der Membran.
HI-Viren sammeln sich vor dem Verlassen der Immunzelle an der Membran. HI-Virus, das sich aus einer Immunzelle herauslöst.
HI-Virus, das sich aus einer Immunzelle herauslöst.Eine unbehandelte HIV-Infektion verläuft in der Regel in mehreren Stadien. Drei bis sechs Wochen nach der Ansteckung kommt es meist zu einer akuten HIV-Infektion. Diese ist durch Fieber, starken Nachtschweiß[45], Abgeschlagenheit, Hautausschläge, orale Ulzerationen oder Arthralgie (Gelenkschmerzen) gekennzeichnet. Wegen der Ähnlichkeit mit grippalen Infektionen bleibt die akute HIV-Infektion meistens unerkannt. Eine frühe Diagnose ist jedoch wichtig: Durch sie können nicht nur weitere Infektionen von Sexualpartnern verhindert werden. Erste Studien an Patienten, die während der akuten HIV-Infektion antiviral behandelt wurden und nach einiger Zeit die Therapie absetzten, zeigten, dass die HIV-spezifische Immunantwort der Patienten gestärkt werden konnte.[46][47] Die akute Infektion dauert selten mehr als vier Wochen an.
In der folgenden, meist mehrjährigen Latenzphase treten keine gravierenden körperlichen Symptome auf. Veränderte Blutwerte und eine schleichende Lipodystrophie bleiben von den HIV-Infizierten oftmals unbemerkt. Danach kommt es vielfach zu ersten Erkrankungen, die auf ein mittelschwer geschwächtes Immunsystem zurückzuführen sind, jedoch noch nicht als AIDS-definierend gelten (CDC Klassifikation B, siehe AIDS).
Zerstörung von CD4-Helferzellen
Im Verlauf einer HIV-Infektion werden unter anderem CD4-Helferzellen kontinuierlich auf verschiedenen Wegen zerstört, was eine Schwächung des Immunsystems bewirkt. Zum einen können infizierte Wirtszellen auf direktem Wege eliminiert werden. Dies geschieht entweder durch Membranschäden an der Zelle, welche durch Ein-/Austritte der Viren verursacht werden, oder durch proapoptotische Eiweiße der HI-Viren sowie zerstörerische Informationshybride aus RNA und DNA. Zum anderen findet eine indirekte Zerstörung infizierter Zellen statt, welche durch gesunde Zellen des Immunsystems als gefährlich erkannt und von ihnen anschließend ausgeschaltet werden. Weiterhin werden auch nichtinfizierte T-Helferzellen als Kollateralschäden durch einen Kontakt mit Proteinen wie p120 zerstört. Diese Proteine entstehen bei der Vermehrung des HI-Virus in der Blutbahn. Im Anschluss an eine akute HIV Infektion und nach erfolgter virusspezifischer Immunantwort ist der Körper in der Regel über einige Jahre in der Lage, die Menge der zerstörten Zellen durch die Produktion neuer Zellen zum größten Teil zu ersetzen.
Ausbildung eines Immundefektes
Bleibt die HIV-Infektion unbehandelt, sinkt die Zahl der CD4 T-Helferzellen kontinuierlich ab, und es kommt im Median neun bis elf Jahre nach der Erstinfektion zu einem schweren Immundefekt (< 200 CD4-Zellen/Mikroliter). Dieser führt in der Regel zu AIDS-definierenden Erkrankungen (CDC Klassifikation C). Zu diesen zählen opportunistische Infektionen, die durch Viren, Bakterien, Pilze oder Parasiten bedingt sind, sowie andere Erkrankungen, wie Kaposi-Sarkom, malignes Lymphom, HIV-Enzephalopathie und das Wasting-Syndrom. Nach individuell unterschiedlicher Zeit führen diese unbehandelt meist zum Tod. Ein schwerer Immundefekt bedeutet jedoch nicht, dass sofort AIDS auftritt. Je länger ein schwerer Immundefekt vorliegt, desto größer ist die Wahrscheinlichkeit, AIDS zu bekommen.
Genetische Faktoren und Resistenz
Die Tatsache, dass Individuen trotz gleicher Infektionsquelle oft sehr unterschiedliche Krankheitsverläufe haben, deutet auf einen starken Einfluss von Wirtsfaktoren auf den Verlauf der Infektion hin. Neben der Ausbildung des Immunsystems scheinen auch einige genetische Faktoren eine Rolle zu spielen. So sind homozygote Individuen mit einem genetischen Defekt am CCR5-Rezeptor (CCR5delta32) weitgehend resistent gegen HIV-Infektionen.[48][49] Dieser Rezeptor dient als Co-Rezeptor bei der Fusion des Virus mit der Wirtszelle. Es wurden nur wenige Individuen gefunden, die eine Infektion trotz Rezeptordefektes haben. Sie infizierten sich mit HI-Viren, die andere Co-Rezeptoren benutzen, wie etwa den CXCR4 Rezeptor auf T-Zellen. Homozygote Genträger dieser Deletion machen etwa ein Prozent der Bevölkerung aus, heterozygote Genträger etwa 20 Prozent. Heterozygote haben zwar deutlich weniger CCR5 Rezeptoren, können sich aber auch mit HIV infizieren und scheinen nach einer Infektion kaum eine längere mittlere Überlebenszeit zu haben.
Der AIDS-Forscher J.J. Bwayo (J.J. Bouyao) untersuchte in Nairobi (Kenia) 600 Prostituierte. Dabei stellte er fest, dass 24 von ihnen offenbar gegen das HI-Virus resistent sind. Der Grund dafür scheint nach Ansicht von Forschern genetisch bedingt zu sein. Offenbar ist eine Gen-Anomalie dafür verantwortlich, die das Virus daran hindert, in die Zellen einzudringen und sich zu verbreiten.
Abgesehen von Mutationen, die eine vollständige Resistenz gegen HIV verleihen, gibt es auch eine Reihe von Genotypen, welche zwar nicht vor einer HIV-Infektion schützen, aber mit einem langsameren Voranschreiten der Krankheit und geringerer Viruslast assoziiert sind. Dabei sind zwei unterschiedliche Mechanismen identifiziert worden:
- Träger gewisser Allele der MHC-I-Proteine, insbesondere HLA-B*5705 und/oder HLA-B*2705, weisen gegenüber anderen Menschen ein langsameres Voranschreiten der Infektion auf. Da MHC I Proteine virale Proteine aus dem Zellinneren binden und so die Infektion einer Zelle anzeigen, wird davon ausgegangen, dass die genannten Varianten in der Lage sind, die Proteine des HIV besonders effizient zu binden. Daher werden HIV-infizierte T-Helferzellen in diesen Individuen besonders schnell von cytotoxischen T-Zellen erkannt und vernichtet.
- Nach einer HIV-Infektion beginnt das Immunsystem mit der Produktion von Antikörpern gegen HIV; aufgrund der schnellen Mutationsrate des HIV bleiben diese aber weitgehend wirkungslos. Einige Menschen produzieren jedoch Antikörper, welche sich gegen eine konstante Region des gp120 richten, was die Infektion verlangsamt. Warum diese Antikörper nur in gewissen Menschen produziert werden, ist unbekannt[50].
Tests auf eine HIV-Infektion
Es gibt verschiedene Möglichkeiten, das Blut, Urin oder auch Gewebe auf die Anwesenheit von HI-Viren oder HIV-Antikörper zu prüfen. Was umgangssprachlich als „Aidstest“ bekannt ist, spürt keinesfalls die Viren selber auf, sondern lediglich die vom Menschen erzeugten Antikörper gegen das eigentliche Virus. Ausnahme sind Tests mit dem PCR-Verfahren, welche mittels Suche nach viralen Nukleinsäuren (Ribonukleinsäure (RNA)) die HI-Viren selbst nachweisen.
Am gebräuchlichsten ist der ELISA-Test und die Western-Blot-Methode. Diese zwei Methoden werden stets gemeinsam verwendet: ELISA ist geeignet, um das Risiko falsch negativer Resultate zu senken; während der genauere Western-Blot-Test falsch positive Resultate vermeidet. ELISA wie Western Blot sind günstige Tests und sie sind ca. zwei bis drei Monate nach einer möglichen Infektion von hoher Genauigkeit, können aber schon zwei bis drei Wochen nach einer vermuteten Ansteckung eingesetzt werden.
Ein positives ELISA-Ergebnis allein ist kein sicherer Befund für eine HIV-Ansteckung; deshalb wird er immer zusammen mit Western Blot angewendet; und es kann vorkommen, dass eine gesunde Person Monate nach einem positiven ELISA-Test wieder einen negativen Befund erhält. Antikörpertests können nach kurz zurückliegenden akuten Erkrankungen, Grippeimpfungen und Allergien falsch-positive Befunde liefern.[51]
Der direkte Nachweis von Virus-Erbgut mittels PCR ist das zuverlässigste, jedoch auch das teuerste Verfahren, welches schon 15 Tage nach einer Ansteckung verlässliche Resultate liefert. Das PCR-Verfahren wird zur Überwachung der antiretroviralen Therapie (HAART), zur Diagnose einer akuten HIV-Infektion und im Blutspendewesen verwendet.
Vorgehensweise beim PCR-Einsatz im Blutspendewesen: Blutspendeproben von Dutzenden von Menschen werden gemischt (engl. Fachbegriff pooling) und dann gemeinsam nach HIV-Erbgut untersucht. Ist das Resultat positiv, werden dann die Blutproben von kleineren und noch kleineren Untergruppen untersucht, um so die Herkunft des gefundenen Viren-Erbgutes auf den einzelnen Spender zurückzuführen.
Siehe auch
- HIV-Impfstoff
- HIV/AIDS in Afrika
- HIV-Prozess in Libyen
- APOBEC3 (Elemente des Immunsystems zur Abwehr von Retrovirus-Infektionen)
- Safer Sex
- Sexualhygiene
- Stiftung Humanitäre Hilfe für durch Blutprodukte HIV-infizierte Personen
- HIV/AIDS in der Ukraine
- Kompetenznetz HIV/AIDS
- WhizzKids United
Literatur
Leitlinien
- S2k-Leitlinie Antiretrovirale Therapie der HIV-Infektion der Deutschen AIDS-Gesellschaft (DAIG). In: AWMF online (Stand 2008)
- S2k-Leitlinie Therapie und Prophylaxe opportunistischer Infektionen bei HIV-infizierten Patienten der Deutschen AIDS-Gesellschaft (DAIG). In: AWMF online (Stand 2011)
- S1-Leitlinie Diagnostik und Behandlung HIV-betroffener Paare mit Kinderwunsch der Deutschen AIDS-Gesellschaft (DAIG). In: AWMF online (Stand 2008)
- S2-Leitlinie Postexpositionelle Prophylaxe der HIV-Infektion der Deutschen AIDS-Gesellschaft (DAIG). In: AWMF online (Stand 2007)
Sonstiges
- Christoph Benn, Sonja Weinreich: HIV und Aids. Eine Krankheit verändert die Welt. Lembeck, Frankfurt am Main 2009, ISBN 978-3-87476-586-2.
- Hans Jäger (Hg.): Entry Inhibitoren. Neue Formen der HIV-Therapie. Springer, Heidelberg 2008, ISBN 978-3-540-78357-2.
- Niko Neye: Humanes Immundefizienz-Virus (HIV). Eine szientometrische Analyse. Freie Universität Berlin 2009. (Dissertation)
Weblinks
 Wiktionary: HIV – Bedeutungserklärungen, Wortherkunft, Synonyme, Übersetzungen
Wiktionary: HIV – Bedeutungserklärungen, Wortherkunft, Synonyme, Übersetzungen Commons: HIV – Sammlung von Bildern, Videos und Audiodateien
Commons: HIV – Sammlung von Bildern, Videos und Audiodateien Wikinews: Humanes Immundefizienz-Virus – in den Nachrichten
Wikinews: Humanes Immundefizienz-Virus – in den Nachrichten- Deutsch
- Informationen des Robert Koch-Instituts zu HIV/AIDS
- Detaillierte und aktuelle Informationen zum Thema HIV auf www.hivbuch.de
- HIV-Diagnostik, auf www.laborlexikon.de
- Kurzstatistiken über HIV und AIDS in Europa (Stand 2004, PDF, 414 KiB)
- Infektionsrisiko bei Oralverkehr
- Stellungnahme des Robert-Koch-Instituts zur Aids-Kritik (PDF, 43 KiB)
- DAGNÄ e. V. (Näheres über HIV unter Fachthemen/HIV-Infektion)
- HIV Factsheet (Stand 2010, PDF, 448 KiB)
- Andere Sprachen
- HIV-Datenbank (englisch)
- HIV InSite (englisch)
- Genom HIV 1
- Genom HIV 2
- Mode de fonctionnement du virus (französisch)
- Intégration et mode expression du virus (französisch)
- Dossier très complet sur le VIH, sa réduplication et les différents tests de dépistages d'Anne Decoster. (französisch)
- The Scientist: 25 Years with HIV – Its progressions, 1981-2006 and beyond (eine Flash-unterstützte Zeitskala) (englisch)
Einzelnachweise
- ↑ AIDS-Epidemie-Update zum Jahresreport 2009 der UNAIDS-Organisation, Stand: Dezember 2009
- ↑ HIV / AIDS in Deutschland – Eckdaten, Epidemiologische Kurzinformation des Robert Koch-Instituts, Stand: Ende 2010
- ↑ Statistiken des Bundesamts für Gesundheit BAG bzgl. „HIV und Aids in der Schweiz: Eckdaten 2006“[1]
- ↑ Zhu, P. et al.: Distribution and three-dimensional structure of AIDS virus envelope spikes. In: Nature. 441, Nr. 7095, 2006, S. 847-852. PMID 16728975.
- ↑ Roux, K.H. and Taylor K. A.: AIDS virus envelope spike structure. In: Curr Opin Struct Biol. 17, Nr. 2, 2007, S. 244-252. PMID 17395457.
- ↑ Liu, J. et al.: Molecular architecture of native HIV-1 gp120 trimers. In: Nature. 455, Nr. 7209, 2008, S. 109-113. PMID 18668044.
- ↑ D.C. Chan, et al.: Core structure of gp41 from the HIV envelope glycoprotein. Cell. 1997 89(2):263-73. PMID 9108481
- ↑ Françoise Barré-Sinoussi, Luc Montagnier, et al.: Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS).Science. 1983 220(4599):868-71. PMID 6189183
- ↑ R.C. Gallo, et al.: Isolation of human T-cell leukemia virus in acquired immune deficiency syndrome (AIDS). Science. 1983 220(4599):865-7. PMID 6601823
- ↑ The Nobel Prize in Physiology or Medicine 2008. In: Internet-Seiten des Nobelpreises. The Nobel Foundation, abgerufen am 18. Dezember 2008 (englisch).
- ↑ Edward Hooper: Aids and the Polio Vaccine. London Review of Books, Vol 25, No. 7 (2003)
- ↑ http://www.uow.edu.au/arts/sts/bmartin/dissent/documents/AIDS/Hooper04/BM6_2.html Edward Hooper. Untruths, misrepresentations and spin: the dubious methods and tactics used by Stanley Plotkin's group in the "Origins of AIDS" debate. 2004
- ↑ Www.aidsorigins.com (Website von Edward Hooper)
- ↑ B. Korber,M. Muldoon,J. Theiler, et al.: Timing the origin of the HIV-1 pandemic. In: Programs and abstracts of the 7th Conference on Retroviruses and Opportunistic Infections. Abstract L5, 30. January 30-2. February 2000.
- ↑ P. Blancou, et al.: Polio vaccine samples not linked to AIDS. In: Nature, 410, S. 1045-1046 (2001)
- ↑ T Zhu, et al.: An African HIV-1 sequence from 1959 and implications for the origin of the epidemic. Nature. 1998 391(6667):594-7. PMID 9468138
Eine fast gleich alte Gewebeprobe aus dem Jahre 1960 wurde 2008 analysiert: Michael Worobey et.al.: Direct evidence of extensive diversity of HIV-1 in Kinshasa by 1960. Nature 455, 2008, S. 661–664, doi:10.1038/nature07390 - ↑ http://www.wissenschaft.de/wissenschaft/news/292503.html Der Werdegang des HI-Virus
- ↑ http://sciencenow.sciencemag.org/cgi/content/full/2008/625/1 E. Pennisi: Revising HIV's History, ScienceNOW Daily News, 25. June 2008
- ↑ http://www.nature.com/nature/journal/v428/n6985/full/428820a.html Worobey M., Mario L. Santiago et al.: Origin of AIDS: Contaminated polio vaccine theory refuted, Nature 428, 820 (22. April 2004); DOI: 10.1038/428820a
- ↑ Sharp, P. M. & Hahn, B. H. (2008): AIDS: prehistory of HIV-1, Nature 455, 605 (2. October 2008); PMID 18833267
- ↑ Worobey, M. et al.(2008): Direct evidence of extensive diversity of HIV-1 in Kinshasa by 1960, Nature 455, 661 (2. October 2008); PMID18833279
- ↑ a b Plantier, J. C. et al.: A new human immunodeficiency virus derived from gorillas. In: Nat. Med.. 15, Nr. 8, 2009, S. 871-872. PMID 19648927.
- ↑ Introduction to HIV types, groups and subtypes. AIDS charity AVERT. Abgerufen am 23. April 2010.
- ↑ a b c d Kirchhoff, F.: "Optimale" Anpassung pandemischer HIV-1-Stämme an den Menschen. In: BIOspektrum. 2, 2010, S. 144-148.
- ↑ Hahn, B. H. et al.: AIDS as a zoonosis: scientific and public health implications. In: Science. 287, Nr. 5453, 2000, S. 607-614. PMID 10649986.
- ↑ Sauter, D. et al.: Tetherin-driven adaptation of Vpu and Nef function and the evolution of pandemic and nonpandemic HIV-1 strains. In: Cell Host Microbe. 6, Nr. 5, 2009, S. 409-421. PMID 19917496.
- ↑ „Optimale“ Anpassung von HIV-1 an den menschlichen Wirt eine Voraussetzung für die effektive Ausbreitung der AIDS Pandemie?. Universität Ulm. Abgerufen am 24. Juni 2010.
- ↑ [2] Bundeszentrale für politische Bildung, 2008
- ↑ CDC: Zahlen zur mittleren Übertragbarkeit von HIV, pro Akt. Abgerufen am 1. Juli 2009.
- ↑ Cornelia Dick-Pfaff: HI-Viren durchdringen auch gesunde Schleimhaut der Scheide. In: wissenschaft-aktuell.de. 17. Dezember 2008, abgerufen am 18. Dezember 2008.
- ↑ CDC: Zahlen zur mittleren Übertragbarkeit von HIV, pro Akt. Abgerufen am 1. Juli 2009.
- ↑ Cornelia Pfaff: Beschneidung schützt vor HIV. In: Internet-Seiten von Bild der Wissenschaft. 26. März 2004, abgerufen am 18. Dezember 2008.
- ↑ Hilgartner, M.: AIDS in the transfusion recipient. In: Pediatr Clin North Am. 38, Nr. 1, 1991, S. 121-131. PMID 1987513.
- ↑ Irja Most: HIV-Infektionsgefahr: Wie sicher sind deutsche Kliniken? Der Tagesspiegel, 29. Februar 2008
- ↑ Deutsch-Österreichische Empfehlungen zur HIV-Therapie in der Schwangerschaft und bei HIV-exponierten Neugeborenen, September 2008[3]
- ↑ CHAT-Survey-Studie
- ↑ Andrea Fischer: Viele holen sich beim Partner HIV. In: Tages-Anzeiger. 2. Mai 2006, abgerufen am 18. Dezember 2008.
- ↑ Deng H et. al.: Identification of a major co-receptor for primary isolates of HIV-1. Nature. 1996 381(6584):661-6 PMID 8649511
- ↑ T. Dragic, et al.: 'HIV-1 entry into CD4+ cells is mediated by the chemokine receptor CC-CKR-5. In: Nature. 1996 381(6584):667-73. PMID 8649512
- ↑ Y.R. Zou, et al.: Function of the chemokine receptor CXCR4 in haematopoiesis and in cerebellar development. Nature. 1998 393(6685):595-9. PMID 9634238
- ↑ C. Winkler, et al.: Genetic restriction of AIDS pathogenesis by an SDF-1 chemokine gene variant. ALIVE Study, Hemophilia Growth and Development Study (HGDS), Multicenter AIDS Cohort Study (MACS), Multicenter Hemophilia Cohort Study (MHCS), San Francisco City Cohort (SFCC) Science. 1998 279(5349):389-93. PMID 9430590
- ↑ Schols D et al.: Inhibition of T-tropic HIV strains by selective antagonization of the chemokine receptor CXCR4. J Exp Med. 1997 186(8):1383-8. PMID 9334378
- ↑ Murakami T et al.: A small molecule CXCR4 inhibitor that blocks T cell line-tropic HIV-1 infection. J Exp Med. 1997 186(8):1389-93. PMID 9334379
- ↑ http://www.hiv-wechselwirkungen.de/media/de/Liste%20der%20antiretroviralen%20Medikamente%20Mai%202008.pdf
- ↑ Armin Schafberger, Holger Sweers: HIV HEUTIGER WISSENSSTAND AIDS. Deutsche AIDS-Hilfe e.V., 2008, S. 7, abgerufen am 10. Januar 2010 (PDF-Datei).
- ↑ Rosenberg ES et al.: Immune control of HIV-1 after early treatment of acute infection. Nature. 2000 407(6803):523-6. PMID 11029005
- ↑ Altfeld M et al.: Cellular immune responses and viral diversity in individuals treated during acute and early HIV-1 infection. J Exp Med. 2001 193(2):169-80. PMID 11148221
- ↑ Dean M et al.: Genetic restriction of HIV-1 infection and progression to AIDS by a deletion allele of the CKR5 structural gene. Hemophilia Growth and Development Study, Multicenter AIDS Cohort Study, Multicenter Hemophilia Cohort Study, San Francisco City Cohort, ALIVE Study. Science. 1996 273(5283):1856-62. Erratum in: Science 1996 Nov 15;274(5290):1069. PMID 8791590
- ↑ Liu R et al.: Homozygous defect in HIV-1 coreceptor accounts for resistance of some multiply-exposed individuals to HIV-1 infection. Cell. 1996 86(3):367-77. PMID 8756719
- ↑ Djordjevic A, Veljkovic M, Antoni S, Sakarellos-Daitsiotis M, Krikorian D, Zevgiti S, Dietrich U, Veljkovic N, Branch DR. The presence of antibodies recognizing a peptide derived from the second conserved region of HIV-1 gp120 correlates with non-progressive HIV infection. Curr HIV Res. 2007 Sep;5(5):443-8. PMID 17896963
- ↑ Simonsen et al: "Multiple false reactions in viral antibody screening assays after influenza vaccination", in American Journal of Epidemiology Vol. 141, No. 11: 1089–1096

Bitte den Hinweis zu Gesundheitsthemen beachten! 

Dieser Artikel wurde in die Liste der lesenswerten Artikel aufgenommen. Kategorien:- Wikipedia:Lesenswert
- Retroviren
- Virusspezies
- Immunologie
- AIDS
- Meldepflichtiger Erreger


{kind=link}
Wikimedia Foundation.