- Dromaeosauridae
-
Dromaeosauridae 
Utahraptor ostrommaysorum, ein Dromaeosaurier aus der Unterkreide Nordamerikas
Zeitraum Mitteljura (Bathonium) bis Oberkreide (Maastrichtium) 167 bis 65,5 Mio. Jahre Fundorte Systematik Dinosaurier (Dinosauria) Echsenbeckendinosaurier (Saurischia) Theropoden (Theropoda) Coelurosauria Deinonychosauria Dromaeosauridae Wissenschaftlicher Name Dromaeosauridae Matthews & Brown, 1922 Die Dromaeosauridae sind eine Gruppe von theropoden Dinosauriern innerhalb der Deinonychosauria. Es handelte sich um kleine bis mittelgroße, zweibeinig laufende Fleischfresser, die wahrscheinlich mit den Vögeln (Aves) nah verwandt waren. Charakteristisch für diese gefiederten Dinosaurier war eine schmale Schnauze, lange, mit gekrümmten Krallen bewehrte Arme, ein versteifter Schwanz sowie eine vergrößerte sichelförmige Kralle am Fuß. Dromaeosauriden wurden in Asien, Europa, Nordamerika, Südamerika, Afrika und Antarktika entdeckt[1] und existierten etwa 100 Millionen Jahre lang, vom Mitteljura bis zur Oberkreide. In der Populärkultur werden Dromaeosauriden auch als „Raptoren“ bezeichnet.
Inhaltsverzeichnis
Merkmale
Größe und Fossilbericht
 Größenvergleich einiger Dromaeosaurier
Größenvergleich einiger Dromaeosaurier

Dromaeosauriden waren kleine bis mittelgroße Tiere. Die Größe reichte von 47 bis 63 Zentimeter Länge und 200 bis 600 Gramm Gewicht bei Microraptor zhaoianus bis über sechs Meter Länge bei Achillobator und Utahraptor. Laut Untersuchungen von Paläontologen um Alan Turner (2007) lassen die generell kleinen Körpermaße der basalen, also am Beginn der Entwicklungslinie stehenden Arten vermuten, dass der gemeinsame Vorfahr aller Dromaeosauriden lediglich etwa 65 Zentimeter lang und 700 Gramm schwer war. Eine Analogie findet sich auch bei der verwandten Gruppe Troodontidae. Die Forscher vermuten daher, dass eine kleine Körpergröße nicht erst von den Vögeln entwickelt wurde, sondern bereits bei den gemeinsamen Vorfahren der Vögel, der Dromaeosauriden und der Troodontiden bestand – eine Gruppe, die auch als Paraves zusammengefasst wird. Im Lauf der Evolution der Dromaeosauriden nahm die Größe mindestens drei Mal unabhängig voneinander drastisch zu: Bei Deinonychus und Unenlagia waren es jeweils mehr als zwei Größenordnungen, während die Größe der Gruppierung Achillobator + Utahraptor um drei Größenordnungen zunahm.[2]
Mit mehr als einem Dutzend Skelettfunden ist Velociraptor der am besten bekannte Vertreter der Dromaeosauridae. Recht vollständiges Skelettmaterial ist auch von Deinonychus, Microraptor und Sinornithosaurus bekannt. Verschiedene Gattungen wie Bambiraptor, Buitreraptor und Tianyuraptor sind jeweils durch ein einziges fast vollständiges Skelett bekannt. Alle anderen Gattungen sind nur durch sehr unvollständige Überreste überliefert.
Schädel

 Schädelschema eines Dromaeosauriden
Schädelschema eines DromaeosauridenDer Schädel war relativ groß und zeigte eine sehr schmale und verlängerte Schnauze. Die von oben betrachtet annähernd dreieckige Form des Schädels ermöglichte nach vorn gerichtete Augen, was auf ein räumliches Sehen hindeutet.[3] Der Hirnschädel war kurz und tief. Wie bei allen Theropoden wies der Schädel mehrere Schädelfenster auf: Die Augenhöhle war groß und rundlich, was auf große Augen hindeutet. Die Nasenöffnungen waren groß und wie ein liegendes Oval geformt. Das Zwischenkieferbein, ein Knochen am vorderen Ende des Oberkiefers, bildete einen langen, nach hinten gerichteten Fortsatz aus, welcher den Oberkiefer von der Nasenöffnung abtrennte – somit wurde die Nasenöffnung lediglich durch das Zwischenkieferbein und das Nasenbein begrenzt. Die vor den Augen gelegene antorbitale Schädelgrube (Fossa) war lang und von drei weiteren Schädelfenstern durchsetzt – dem Antorbitalfenster, dem Maxilliarfenster und dem Promaxilliarfenster. Der Unterkiefer zeigte ein dünnes und verlängertes Mandibularfenster. Am hinteren Ende der Schädeldecke bildeten das paarige Scheitelbein und das Schuppenbein einen scharfen, nach oben gerichteten Schädelkamm.
Der Schädel zeigt verschiedene einzigartige Merkmale, durch welche die Gruppe von anderen Gruppen abgegrenzt wird (Autapomorphien). So war das paarige Stirnbein „T“-förmig und zeigte eine sinusförmige Begrenzung des Subratemporalfensters, ein Schädelfenster hinter den Augen. Weitere einzigartige Merkmale des Schädels sind ein überhängender Fortsatz des Schuppenbeins, der sich bis über den Kopf des Quadratbeins erstreckte, sowie ein flügelartiger Fortsatz des Quadratbeins, der das Quadratojugale auf der Oberseite (dorsal) berührte.
Ein vollständiger Gaumen wurde bisher bei keinem Exemplar gefunden, obwohl die meisten Knochen des Gaumens bekannt sind. Erst jüngere Studien haben ergeben, dass Dromaeosauriden einen sekundären Gaumen (eine knöcherne Abtrennung zwischen Mund- und Nasenraum) aufwiesen. Das Zwischenkieferbein zeigte einen fast quadratischen Hauptkörper und vier schwach gekrümmte Zähne, während im Oberkiefer 9 bis 15 Zähne steckten. Die Zahnreihe des Oberkiefers endete noch vor dem Beginn der Augenhöhle. Der Unterkiefer war bei Velociraptor lang und dünn, bei Deinonychus und Dromaeosaurus aber tiefer und robuster. Im Dentale, dem zahntragenden Teil des Unterkiefers, steckten 11 bis 16 Zähne. Das Surangulare, ein Kieferknochen hinter dem Dentale, zeigte ein markantes Loch (Foramen). Die Zähne waren groß, leicht nach hinten gebogen und außer bei Buitreraptor[3] auf der Rückseite gesägt. Charakteristisch für Dromaeosauriden war, dass die Sägung zur Zahnspitze hin größer wurde.
Rumpfskelett und Gliedmaßen
 Skelett eines Deinonychus
Skelett eines Deinonychus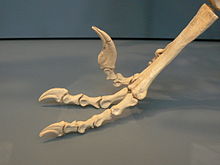 Fußskelett eines Dromaeosaurus
Fußskelett eines DromaeosaurusNahezu komplett erhaltene Wirbelsäulen sind lediglich von Velociraptor bekannt, welcher 9 Hals-, 12 Rücken-, 5 bis 6 verschmolzene Kreuzbein-, und mindestens 43 Schwanzwirbel aufwies. Der Hals war beim lebenden Tier deutlich „S“-förmig gebogen. Die Wirbelzentren der meisten Hals- und Rückenwirbel waren kürzer und breiter als bei anderen Coelurosauriern. Ein einzigartiges Merkmal der Dromaeosauriden waren die vergrößerten Parapophysen der Rückenwirbel, Fortsätze auf der Unterseite von Wirbeln. Der Schwanz war lang und durch stark verlängerte, stangenartige Präzygapophysen (mechanische Verbindungselemente der Wirbel) verstärkt. Verlängerte Präzygapophysen wurden ab dem elften Schwanzwirbel ausgebildet und verliefen in verwobenen Bündeln entlang der Wirbelbögen nach vorne. Ein einziger dieser Wirbelauswüchse konnte bis zu zehn weitere Wirbel verbinden. Analog zu den Präzygapophysen bildeten die Chevronknochen an der Unterseite der Schwanzwirbel ebenfalls lange, nach vorn gerichtete Fortsätze, die jeweils mehrere Wirbel überragten und in verwobenen Bündeln angeordnet waren. In seiner Beschreibung des Deinonychus vermutet John Ostrom (1969), diese Verbindungselemente hätten den Schwanz derart versteift, so dass dieser nur am Schwanzansatz beweglich war. Ein neuerer Fund eines Velociraptor zeigt jedoch einen Schwanz im anatomischen Verbund, der horizontal „S“-förmig gebogen war, ohne dass die Anordnung der Präzygapophysen beeinträchtigt wurde. Dies weist auf eine beträchtliche seitliche Beweglichkeit des Schwanzes hin.[4]
Die Arme gehörten zu den im Verhältnis längsten aller Theropoden und machten bei Velociraptor 65 Prozent und bei Sinornithosaurus 80 Prozent der Hinterbeinlänge aus. Der Unterarm macht 75 Prozent der Länge des Oberarms bei Deinonychus und 70 Prozent bei Velociraptor aus. Die Hand war lang und machte bei Deinonychus 40 Prozent der Armlänge aus. Jeder der drei Strahlen der Hand endete in großen, gekrümmten Klauen. Der erste Strahl war der kürzeste, während der zweite Strahl der längste war. Der dritte Strahl war etwas kürzer aber deutlich dünner als der zweite Strahl. Das zweite und das dritte Fingerglied (Phalanges) des dritten Strahls waren extrem verkürzt – ähnlich wie bei Archaeopteryx, aber anders als bei anderen Coelurosauriern. Biomechanische Studien haben ergeben, dass Dromaeosauriden unter Zuhilfenahme beider Hände Objekte greifen konnten. Bambiraptor konnte den ersten und dritten Finger jeden anderen Finger gegenüberstellen und somit Gegenstände mit nur einer Hand greifen – derartige opponierbare Finger sind von keinem anderen Theropoden bekannt.[5]
Das Schambein war lang und nach hinten geneigt, bei den basalen Unenlagiinen jedoch vertikal ausgerichtet.[3] Der Oberschenkel war 10 bis 20 Prozent kürzer als der Unterschenkel, außer bei Achillobator, der einen längeren Oberschenkel als Unterschenkel zeigte. Der Mittelfuß war kurz. Die Mittelfußknochen II, III und IV waren gut entwickelt und etwa gleich groß, während der erste und der fünfte Mittelfußknochen reduziert waren. Die zweite Zehe zeigte eine vergrößerte, sichelartig gebogene Klaue. Die Größe dieser „Sichelkralle“ variierte bei verschiedenen Gattungen stark – während sie beispielsweise bei Deinonychus und Velociraptor extrem vergrößert war, war sie bei Adasaurus eher klein. Zumindest einige Dromaeosauriden trugen den zweiten Zeh mit der vergrößerten Klaue während der Fortbewegung über dem Boden, waren also funktional zweizehig. Hierauf weisen zweizehige Fußspuren sowie viele guterhaltene Skelette hin, bei denen die Sichelkralle in einer aufgerichteten Position erhalten ist. Fossile Fußspuren weisen des Weiteren darauf hin, dass die Fußbasis von einem fleischigen Polster unterstützt wurde, anders als bei anderen Theropoden.[6]
Paläobiologie
Funktion der Sichelkralle
 Die berühmten „kämpfenden Dinosaurier“
Die berühmten „kämpfenden Dinosaurier“Bereits Ostrom (1969) spekuliert bei seiner Beschreibung des Deinonychus („schreckliche Klaue“), die vergrößerte „Sichelkralle“ am zweiten Zeh könnte zur Tötung von Beutetieren gedient haben. Nach Ostroms Vorstellung konnte Deinonychus große ornithopode Dinosaurier mit seinen langen Armen festhalten, um deren Bäuche mit den Fußkrallen aufzuschlitzen. Zur Untermauerung dieser Hypothese weist Ostrom auf Berichte über heutige Laufvögel wie Kasuare oder Strauße hin, die mit ihren verlängerten Krallen am zweiten (inneren) Zeh Menschen aufgeschlitzt haben.[7]
Der 1971 in der Mongolei gemachte spektakuläre Fund der „kämpfenden Dinosaurier“ unterstützt die Idee, dass die „Sichelkralle“ zur Tötung gedient haben könnte. Dieser berühmte Fund zeigt die Skelette von Velociraptor und dem Ceratopsia Protoceratops, welche sich offenbar mitten in einem Kampf befanden, als sie starben: Während das Velociraptor-Exemplar – auf der rechten Seite liegend – den linken Fuß mit nach unten zeigender Sichelkralle in die Halsregion des Protoceratops ausstreckt, scheint der rechte Fuß unter dem liegenden Körper des Protoceratops-Exemplars eingeschlossen zu sein. Die linke Hand von Velociraptor liegt direkt hinter dem Wangenhorn von Protoceratops, während letzterer den rechten Unterarm von Velociraptor mit seinem Schnabel festhält. In dieser Position könnten die kämpfenden Tiere von einem Sandsturm überrascht und begraben worden sein – auch ist möglich, dass die bereits toten Tiere von Flugsand bedeckt wurden. Da die Sichelkralle zum Hals hin ausgestreckt war, vermutet Kenneth Carpenter (1998), dass sie nicht zum Aufschlitzen, sondern zum Durchstechen der Halsadern oder der Luftröhre verwendet wurde. So hätte die Kralle die besonders dicke Bauchhaut und deren Muskeln nicht aufschlitzen können; auch war die vermutete Schneidekante der Kralle abgerundet, und der Keratinüberzug beim lebenden Tier wäre wahrscheinlich weniger scharf gewesen als ein stumpfes Messer. Auf die von Ostrom aufgeführten Berichte über Laufvögel, die mit ihren verlängerten Fußkrallen Tiere und Menschen aufschlitzen können, schreibt Carpenter, dass es sich dabei um sehr seltene Ereignisse handelt und dass das Aufschlitzen durch das große Gewicht der Laufvögel zustande kommt, nicht durch die Schneidewirkung der Kralle selbst – dies lässt es unwahrscheinlich erscheinen, dass die leichtgewichtigen Dromaeosauriden mit ihrer Sichelkralle eine Schneidewirkung vollziehen konnten.[8]
Forscher um Phillip Manning (2005) vermuten, dass die Kralle wie ein Steigeisen fungierte, mit dem sich die Tiere an großen Beutetieren festhalten konnten. So haben Versuche mit einem hydraulischen Robotermodell des Fußes von Deinonychus ergeben, dass die Kralle kleine und rundliche Einstichlöcher erzeugte und dass das Gewebe unterhalb der Einstichlöcher stark zusammengeschoben wurde, was der Kralle zusätzlichen Halt verschaffte. Nach der Vorstellung dieser Forscher haben Dromaeosauriden große Beutetiere angesprungen und konnten mit ihren gesägten Zähnen viele Verletzungen verursachen, während sie mit ihren Fuß- und Armkrallen fest am Beutetier verankert waren.[9]
Eine neuere Studie unter Leitung von Manning (2009) kommt zu dem Schluss, dass die Sichelkralle und die anderen stark gekrümmten Krallen der Hände und Füße nicht nur zum Beutefang, sondern auch ideal zum Klettern geeignet waren. Dazu röntgten die Forscher eine Handklaue von Velociraptor, erstellten daraus ein dreidimensionales Kontourdiagramm und verglichen es mit Krallen des heutigen Uhus, um die Verteilung von Belastungen auf die Klaue zu untersuchen. Wie frühere Studien an heutigen Säugetieren und Vögeln gezeigt haben, lassen sich baumbewohnende Tiere anhand der Krümmung ihrer Krallen von bodenbewohnenden Tieren unterscheiden. Die starke Krümmung der Krallen von Dromaeosauriden ähnelt derer heutiger baumbewohnender Tiere, was nach Ansicht der Forscher auf eine baumbewohnende Lebensweise hinweist. Des Weiteren vermuten die Forscher, dass der zweite, die Sichelkralle tragende Zeh dank modifizierter Sehnen beim Greifen mechanisch sperrte, und sich erst beim Heben des Fußes wieder lösen ließ. Diese Anpassung findet sich bei heutigen baumbewohnenden Vögeln und ermöglicht beispielsweise das Schlafen auf Zweigen ohne herunterzufallen. Bei Dromaeosauriden könnte eine derartige Anpassung eine Hilfe beim Klettern und beim Festkrallen an großen Beutetieren gewesen sein.[10]
Rudelleben und Fußspuren
 Velociraptor hinterlässt Fußspuren im Sand
Velociraptor hinterlässt Fußspuren im SandIn Montana wurden Knochen von mindestens drei Deinonychus-Individuen zusammen mit den Überresten des pflanzenfressenden Tenontosaurus entdeckt. Ostrom und Maxwell (1995) vermuten, dass eine Gruppe von Deinonychus den wesentlich größeren Pflanzenfresser angegriffen hat und dass die gefundenen Deinonychus-Knochen von Tieren stammen, die im darauffolgenden Kampf umgekommen sind. Zähne von Deinonychus werden gelegentlich zusammen mit Überresten des Tenontosaurus gefunden, woraus diese Forscher schließen, dass Tenontosaurus das wichtigste Beutetier von Deinonychus war.[11] Carpenter (1998) merkt jedoch an, dass es ebenso wahrscheinlich ist, dass Deinonychus lediglich als Aasfresser an den Kadavern der Tenontosaurus gefressen hat.[8] Brinkman und Roach (2007) vermuten, dass Deinonychus und andere Theropoden keine Rudeltiere waren, sondern sich ähnlich wie heutige Krokodile als konkurrierende Einzelgänger an Kadavern versammelten. So fanden die Forscher bei Deinonychus Hinweise auf Kannibalismus.[12]
Zweizehige Fußspuren aus Shandong (China), die wahrscheinlich von Dromaeosauriden stammen, weisen darauf hin, dass sich diese Tiere zumindest gelegentlich zu Gruppen zusammengeschlossen haben. Die Fundstelle zeigt sechs Fährtenfolgen, die im engen Abstand parallel zueinander in dieselbe Richtung verlaufen. Der Abstand zwischen den einzelnen Trittsiegeln ist bei allen Fährtenfolgen etwa gleich, was anzeigt, dass alle Individuen mit gleicher Geschwindigkeit gegangen sind. Das Fehlen von Trockenrissen an den Spuren weist darauf hin, dass die Spuren schon bald nach dem Entstehen von Sedimenten bedeckt wurden. Li und Kollegen (2008) merken daher an, dass es sehr unwahrscheinlich sei, dass die Spuren nicht von einer Gruppe, sondern nacheinander von unabhängigen Individuen erzeugt wurden.[6]
Die Spuren stammen vermutlich von einem großen, über fünf Meter langen, mit Achillobator vergleichbaren Dromaeosauriden, worauf die 26 bis 28 Zentimeter langen Trittsiegel hindeuten. Sie wurden als neuer Spurentyp (sogenannte Ichnospezies) beschrieben und Dromaeopodus genannt. Derartige Ichnospezies sind nicht Teil der klassischen biologischen Systematik, sondern gehören zu einem Klassifikationssystem speziell für fossile Fußspuren, das komplett von der klassischen Systematik abgekoppelt ist. Eine weitere, wahrscheinlich den Dromaeosauriden zuzuschreibende Ichnospezies ist Velociraptorichnus, die kleinere, etwa zehn Zentimeter lange Abdrücke zeigt. Zweizehige Fußspuren, die zu Dromaeosauriden gehören könnten, sind sehr selten und lediglich aus China (Gansu, Sichuan und Shandong), Südkorea und den Vereinigten Staaten (Utah) bekannt.[6]
Weitere Hinweise auf die Ernährung und Giftigkeit
Obwohl oft vermutet wird, dass Dromaeosauriden große Beutetiere wie Ornithopoden jagen konnten, ernährten sie sich wahrscheinlich wie heutige Raubtiere sowohl von kleinen als auch von großen Beutetieren.[5] Das Skelett eines azhdarchiden Flugsauriers (Pterosauria) aus dem Dinosaurier-Provinzpark von Alberta (Kanada) weist Bissspuren sowie einen abgebrochenen Zahn des Dromaeosauriden Saurornitholestes langstoni auf, der im Schienbein des Fundes steckt. Ob der Dromaeosauride den Flugsaurier erlegt hat ist jedoch fraglich, da der Flugsaurier mit einer geschätzten Flügelspannweite von sechs Metern um ein Vielfaches größer war als der weniger als zwei Meter große Dromaeosauride. In diesem Fall war der Dromaeosauride daher wahrscheinlich ein Aasfresser.[13]
Eine jüngere Studie aus 2009 fand bei Sinornithosaurus, einem kleinen gefiederten Dromaeosauriden der Jehol-Gruppe, Hinweise auf einen giftigen Biss. So zeigt diese Gattung neben einer ungewöhnlichen Bezahnung gekerbte Zähne, eine Höhlung, welche vermutlich eine Giftdrüse repräsentierte, sowie einen Kanal, der von dieser Giftdrüse zur Zahnbasis führte. Diese Merkmale sind analog zu heutigen giftigen Echsen. Die Forscher vermuten, dass Sinornithosaurus und verwandte Dromaeosauriden Jagd auf Vögel gemacht haben könnten.[14]
Federn
 Fossil eines Sinornithosaurus, der erste Nachweis von Federn bei Dromaeosauriden
Fossil eines Sinornithosaurus, der erste Nachweis von Federn bei DromaeosauridenDer erste direkte Nachweis von Federn bei Dromaeosauriden gelang 1999 mit der Beschreibung von Sinornithosaurus. Dieses Skelett zeigt an verschiedenen Körperpartien Bereiche mit ungefähr vier Zentimeter langen Strukturen, deren Anatomie jedoch nicht erkennbar ist.[15] Seit diesem Fund wurden verschiedene weitere gefiederte Dromaeosauriden entdeckt. Das 2000 beschriebene Typusexemplar von Microraptor zahoianus zeigt Abdrücke, welche vermutlich von Federschäften (Rachis) stammen, was auf echte Konturfedern schließen lässt.[16] Ein weiteres, 2001 beschriebenes Fossil, das möglicherweise zu Sinornithosaurus gehört, zeigt Federn an allen Körperpartien außer dem unteren Abschnitt der Beine.[17] Weitere Fossilien von Microraptor, insbesondere die Funde von Microraptor gui, lassen auf Arm- und Beinschwingen schließen, die ähnlich wie die Flügel heutiger Vögel aufgebaut waren. Einige dieser Schwungfedern weisen asymmetrische Federfahnen auf, was auf eine aerodynamische Funktion schließen lässt. Manche Schwungfedern konnten mehr als doppelt so lang werden wie der Oberschenkelknochen.[18]
Sämtliche gefiederte Dromaeosauriden stammen aus der Jehol-Gruppe in Liaoning (China), einer bedeutenden Fossillagerstätte. Vulkanische Aschen trugen zur Zeit dieser Ablagerungen entscheidend zur Entstehung von außergewöhnlich vollständigen Fossilien bei, so dass auch Federn erhalten blieben, die andernorts nur sehr selten fossil überliefert sind. Direkte Hinweise auf Federn außerhalb der Jehol-Fauna finden sich bei Velociraptor und Rahonavis: Bei diesen Gattungen wurden Ansatzstellen für Federn (englisch quill knobs) an der Elle des Unterarms gefunden, was auf lange Armfedern schließen lässt.[19][20] Federn dienten ursprünglich möglicherweise als Körperisolierung.[21] Spezielle Kopffedern bei Microraptor könnten zur Zurschaustellung gedient haben.[18] Lange Armfedern, wie sie bei Velociraptor vermutet werden, könnten darüber hinaus auch eine isolierende Funktion bei Bebrütung der Eier gehabt haben, worauf Funde des Oviraptorosauriers Citipati hindeuten, die dieses Tier in einer vogelähnlichen Brutposition über einem Nest zeigen.
Flug
 Fossil eines Microraptor gui mit Abdrücken gefiederter Flügel
Fossil eines Microraptor gui mit Abdrücken gefiederter FlügelBasale Dromaeosauriden zeigen sehr ähnliche Anpassungen an den Flug wie der Urvogel Archaeopteryx. Unterschiede im Flugapparat finden sich lediglich in den Proportionen der Arme, die beispielsweise bei Archaeopteryx länger waren als bei Dromaeosauriden.[18][15] Die Fähigkeit zum aktiven Flug oder zum Gleitflug wurde für mindestens zwei Spezies vorgeschlagen. Microraptor gui zeigte Flügel mit asymmetrischen Schwungfedern an Armen und Beinen, die er wahrscheinlich zum Gleitflug von Baum zu Baum einsetzen konnte. Xu und Mitarbeiter vermuten bei ihrer Beschreibung dieses Tieres, dass die langen Beinfedern ein schnelles Fortbewegen auf dem Boden unmöglich gemacht hätten und dass basale Dromaeosauriden somit baumbewohnend (arboreal) gewesen wären. Nach diesen Forschern konnte Microraptor gui seine Beinflügel analog zu den Armflügeln seitlich ausstrecken, um so mit einem vorderen und einen hinteren Paar Flügeln zu gleiten.[18] In einer neueren Studie vermuten Chatterjee und Templin jedoch eine gestufte doppeldeckerähnliche Anordnung der Flügel: Nach diesen Forschern waren die Beine beim Gleitflug nicht horizontal, sondern vertikal entlang der Sagittalebene ausgerichtet und „Z“-förmig eingezogen, während die Schwungfedern seitlich ausgerichtet waren. Die Beinflügel wurden wahrscheinlich tiefer als die Armflügel gehalten und konnten somit unter die Armflügel ausgestreckt werden. Vermutlich konnte Microraptor mit einem wellenförmigen Gleitflug Distanzen von über 40 Metern überbrücken, bei einer Geschwindigkeit von 9 bis 15 m/s. Die Forscher halten einen begrenzten aktiven Flug bei Microraptor theoretisch für möglich. Microraptor gui wird als möglicher Hinweis darauf gedeutet, dass es in der Entwicklung des Vogelflugs ein Zwischenstadium mit vier Flügeln gab, bis sich die Konfiguration mit zwei Flügeln durchsetzte, wie sie bereits Archaeopteryx zeigte. Alternativ könnte es sich bei der vierflügeligen Konfiguration von Microraptor aber auch um eine evolutionäre Sackgasse gehandelt haben.[22]
Für einen anderen basalen Dromaeosauriden der Jehol-Gruppe, Cryptovolans pauli, wurde die Fähigkeit zum aktiven Flug vorgeschlagen,[23] wobei Cryptovolans wahrscheinlich mit Microraptor identisch ist.[24] Rahonavis aus der Oberkreide Madagaskars könnte ebenfalls zum aktiven Flug fähig gewesen sein, worauf deutliche Anpassungen hinweisen – insbesondere ähnelt die Struktur des Schultergürtels dieser Spezies mehr derjenigen heutiger Vögel als der von Archaeopteryx. Chiappe und Dyke (2006) vermuten, dass Rahonavis ähnlich wie Archaeopteryx ein schwächerer und weniger wendiger Flieger war als die meisten heute lebenden Vögel.[20] Rahonavis gilt traditionell als echter Vogel und wird erst in jüngeren Studien innerhalb der Dromaeosauridae eingeordnet.[3][25]
Paläoökologie, Ursprung und Paläobiogeographie
Dromaeosauriden sind aus einer großen Bandbreite verschiedener Habitate bekannt. Während die Überreste von Deinonychus, Dromaeosaurus und Saurornitholestes aus fluviatilen, in Flüssen abgelagerten Sedimenten stammen, wurde Velociraptor in sandigen Sedimenten entdeckt, die auf ein trockenes, wüstenartiges Habitat hindeuten. Obwohl sie traditionell als flinke, terrestrische Räuber dargestellt werden, vermutet bereits Chatterjee (1997) wegen des nach hinten gerichteten Schambeins und wegen Merkmale an der Hand, dass einige Dromaeosauriden baumbewohnend waren. Kürzlich wurde mit der Beschreibung des Microraptorinen Hesperonychus aus der Dinosaurier-Park-Formation von Alberta (Kanada) bekannt, dass Microraptorinen nicht nur in der Kreide Asiens, sondern auch in der Kreide Nordamerikas die Nische der kleinen Prädatoren besetzt haben könnten.[26] Zahnfunde aus dem Oberjura Deutschlands (Oker, Niedersachsen) sowie aus Portugal weisen darauf hin, dass Dromaeosauriden auf dem europäischen Archipel zu dieser Zeit verbreitet waren.[27]
Ostrom (1990) vermutet, dass die Dromaeosauridae während der Unterkreide (Barremium) in Nordamerika entstanden sind und sich dann über Asien und Europa ausgebreitet haben. Jedenfalls wurden die Dromaeosauridae schon 1992 aus dem mittleren Jura (Bathonium) nachgewiesen – ein Zahnfund aus der englischen Grafschaft Gloucestershire ist der älteste bisher entdeckte Nachweis dieser Gruppe.[28] Norell und Makovicky (2004) halten derweil einen Ursprung der Gruppe in Asien für wahrscheinlich, da die meisten und insbesondere die basalsten Gattungen aus diesem Kontinent stammen. Einige neuere Studien gehen jedoch davon aus, dass die Dromaeosauride zu einer Zeit entstand, als noch alle Landmassen zum Superkontinent Pangaea vereint waren. Die Dromaeosauriden könnten somit wie andere Dinosauriergruppen kosmopolitisch über Pangaea verbreitet gewesen sein, bevor dieser im mittleren oder oberen Jura in den Nordkontinent Laurasia und den Südkontinent Gondwana zerbrach. Diese Studien sehen Hinweise bei einigen wenigen Dromaeosauriden aus Gondwana, die allesamt zu einer eigenständigen, basalen Gruppe zusammengefasst werden können, den Unenlagiinen. So vermuten Makovicky und Kollegen (2005), dass sich die Unenlagiinen seit der Trennung Gondwanas von Laurasia unabhängig von den anderen Dromaeosauriden der Nordkontinente entwickelt haben.[3][1] Turner und Kollegen (2007) widersprechen dieser Hypothese einer monophyletischen, auf die Südhalbkugel beschränkten Gruppe, und verweisen auf Shanag, den sie als einen Unenlagiinen aus der Nordhalbkugel sehen.[2] Longrich und Currie (2009) führen Shanag jedoch als Microraptorinen.[26]
Systematik
Äußere Systematik und Verwandtschaft mit Vögeln
Nach der verbreiteten Ansicht waren die Troodontiden die nächsten Verwandten der Dromaeosauriden. Zusammen bilden diese beiden Gruppen die Deinonychosauria, eine Gruppe innerhalb der maniraptoren Theropoden. Ostrom (1973) erkannte als Erster deutliche Ähnlichkeiten zwischen Dromaeosauriden und Vögeln und schlussfolgerte in späteren Arbeiten eine nahe Verwandtschaft dieser beiden Gruppen – eine Vermutung, die seitdem von vielen Studien bestätigt wurde. Die genauen Verwandtschaftsbeziehungen zwischen Dromaeosauriden und Vögeln sind jedoch noch immer Gegenstand vieler Debatten. Die meisten Forscher gehen davon aus, dass die Vögel die Schwestergruppe der Deinonychosauria bilden – diese beiden Gruppen werden auch als Paraves zusammengefasst.[29][2]
Norell und Kollegen (2001) sehen die Dromaeosauriden als näher mit den Vögeln verwandt als mit den Troodontiden – sie vermuten sogar, dass die Dromaeosauridae als paraphyletische Gruppe die Vögel mit einschließen, dass also alle Vögel von Dromaeosauriden abstammen.[17] Larry Martin (2004) vermutet, dass die gesamte Gruppe der Maniraptora einschließlich der Dromaeosauriden keine Dinosaurier, sondern sekundär flugunfähige Vögel waren. Martin zufolge spalteten sich die Vögel während der Trias von basalen, vierfüßigen Archosauriern ab.[30] Auch Feduccia und Kollegen (2005) widersprechen der verbreiteten Ansicht einer Abstammung der Vögel von den Dinosauriern. Nach diesen Forschern muss zumindest die Dromaeosauriden-Untergruppe Microraptorinae innerhalb der Vögel eingeordnet werden.[24]
Innere Systematik
Innerhalb der Dromaeosauridae werden bis zu sechs Gruppen unterschieden. Einige aus den südlichen Kontinenten stammende Gattungen bilden die wenig bekannte Gruppe Unenlagiinae, die häufig als die basalste (urtümlichste) Gruppe betrachtet wird.[31] Ebenfalls basal waren die Microraptorinae, die sehr kleine und vermutlich baumbewohnende Dromaeosauriden umfasst. Sämtliche Dromaeosauriden, von denen Federabdrücke erhalten sind, stammen aus dieser Gruppe. Die fortgeschritteneren Gruppen der Dromaeosauriden werden von Longrich und Currie (2009) auch als Eudromaeosauria zusammengefasst:[26] Während die Velociraptorinae Gattungen wie Velociraptor und Deinonychus umfasst, werden der Dromaeosaurinae Dromaeosaurus sowie die großwüchsigen Gattungen Utahraptor und Achillobator zugeschrieben. Saurornitholestes wird manchmal zusammen mit Bambiraptor und Atrociraptor in eine eigene Gruppe eingeordnet, die Saurornitholestinae.
Die folgende Gattungsliste basiert auf Longrich und Currie (2009):[26]
- Dromaeosauridae
- Mahakala
- Dromaeosaurinae
- Microraptorinae
- Saurornitholestinae
- Unenlagiinae
- Velociraptorinae
- Status unklar (nach Norell und Makovicky, 2004)[34]
- Pyroraptor
- Unquillosaurus
- Variraptor
- Dromaeosauroides
- Luanchuanraptor
Dromaeosauridae und Menschen
Entdeckungs- und Forschungsgeschichte
 Ähnlichkeiten der Hände von Deinonychus (links) und Archaeopteryx (rechts) ließen Ostrom (1976) vermuten, dass Vögel von Dinosauriern abstammen.
Ähnlichkeiten der Hände von Deinonychus (links) und Archaeopteryx (rechts) ließen Ostrom (1976) vermuten, dass Vögel von Dinosauriern abstammen.1922 beschrieben Matthew und Barnum Brown mit Dromaeosaurus den ersten Dromaeosauriden – basierend auf einem teilweise erhaltenen Schädel und Fußknochen, die Brown 1914 in der Dinosaurier-Park-Formation in Alberta entdeckte. Diese Forscher schrieben Dromaeosaurus der Deinodontidae (heute Tyrannosauridae) zu, stellten ihn innerhalb dieser Gruppe jedoch in eine eigene Unterfamilie, die Dromaeosaurinae.[35] Der Name Dromaeosauridae bedeutet soviel wie „laufende Echsen“ (gr.: dromaios = „rennen“, sauros = „Echse“) und soll auf Tiere hinweisen, die im Gegensatz zu anderen Deinodontiden klein und leichtgebaut waren.[36] Bereits zwei Jahre später (1924) beschrieb Henry Fairfield Osborn einen nahezu kompletten Schädel und ein Fingerglied als Velociraptor. Osborn hielt Velociraptor jedoch für einen Megalosauriden.[37]
Ostrom (1969) bot mit seiner Beschreibung des Deinonychus erstmals eine detailliertere Beschreibung, auch des Postkraniums. Er stellte sich Deinonychus als aktiven, schnellen Räuber vor und widersprach damit der damals verbreiteten Vorstellung der Dinosaurier als langsame, unbeholfene Kreaturen. 1976 bemerkte Ostrom deutliche Ähnlichkeiten der Handskelette von Deinonychus und des Urvogels Archaeopteryx und schloss daraus, dass Vögel von theropoden Dinosauriern abstammen.[38] 1999 wurde mit Sinornithosaurus der erste von mehreren gefiederten Dromaeosauriden aus der Jehol-Gruppe von China geborgen. Im selben Jahr wurde im National Geographic Magazine die Entdeckung von „Archaeoraptor“ bekanntgegeben und als „fehlendes Bindeglied zwischen Dinosaurier und Vögel“ gekürt. Später konnte dieses Fossil als geschickte Fälschung entlarvt werden, die aus den Überresten verschiedener Tiere zusammengesetzt war – so gehörte der Großteil des Skeletts zu dem kreidezeitlichen Vogel Yanornis, während der Schwanz zu dem im Jahr 2000 beschriebenen Dromaeosauriden Microraptor gehörte.[39] Jüngere Entdeckungen sind die Microraptorinen Hesperonychus und Tianyuraptor, die beide 2009 beschrieben wurden.
In der Populärkultur
Einige Vertreter der Dromaeosauride, wie Deinonychus, Velociraptor und Utahraptor, erscheinen regelmäßig in populären Darstellungen über Dinosaurier. Velociraptor nimmt in dem 1990 erschienen Science-Fiction-Roman „Jurassic Park“ (deutscher Titel auch „DinoPark“) von Michael Crichton sowie in Steven Spielbergs gleichnamiger Verfilmung von 1993, eines der kommerziell erfolgreichsten Werke der Filmgeschichte, einen bedeutenden Teil der Handlung ein und zählt heute zu den populärsten Dinosauriern. Spielberg war jedoch von der geringen Körpergröße von Velociraptor enttäuscht und stellte dieses Tier im Film stark vergrößert dar.[40] Ein weiteres Beispiel ist der Roman „Raptor Red“ (1995) des Paläontologen Robert Bakker, der aus der Sicht eines weiblichen Utahraptors geschrieben ist.
Einzelnachweise
- ↑ a b J. A. Case, J. E. Martin, M. Reguero: A dromaeosaur from the Maastrichtian of James Ross Island and the Late Cretaceous Antarctic dinosaur fauna. In: A. Cooper, C. Raymond, I. E. Team (Hrsg.): Antarctica: a Keystone in a Changing World – Online Proceedings for the Tenth International Symposium on Antarctic Earth Sciences. U.S. Geological Survey Open-File Report 2007–1047, SRP 083. U.S. Geological Survey, Washington 2007, S. 1–4.
- ↑ a b c Alan H. Turner, Diego Pol, Julia A. Clarke, Gregory M. Erickson, Mark Norell: A basal dromaeosaurid and size evolution preceding avian flight. In: Science. 317, 2007, S. 1378–1381, doi:10.1126/science.1144066.
- ↑ a b c d e Peter J. Makovicky, Sebastián, Agnolín Apesteguía, L. Federico: The earliest dromaeosaurid theropod from South America. In: Nature. 437, 2005, S. 1007–1011, doi:10.1038/nature03996.
- ↑ Mark A. Norell, Peter J. Makovicky: Important features of the dromaeosaurid skeleton II: information from newly collected specimens of Velociraptor mongoliensis. In: American Museum Novitates. 3282, 1999, S. 1–45].
- ↑ a b Phil Senter: Comparison of forelimb function between Deinonychus and Bambiraptor (Theropoda: Dromaeosauridae). In: Journal of Vertebrate Paleontology. 26, Nr. 4, 2006, S. 897–906, doi:10.1671/0272-4634(2006)26[897:COFFBD]2.0.CO;2.
- ↑ a b c Rihui Li u. a.: Behavioral and faunal implications of Early Cretaceous deinonychosaur trackways from China. In: Naturwissenschaften. 95, Nr. 3, 2008, doi:10.1007/s00114-007-0310-7.
- ↑ John H. Ostrom: Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana. Peabody Museum of Natural History, New Haven 1969, S. 1–165 (Peabody Museum of Natural History Bulletin. Band 30).
- ↑ a b Ken Carpenter: Evidence of predatory behavior by theropod dinosaurs. In: Gaia. 15, 1998, S. 135–144.
- ↑ Phillip L Manning u. a.: Dinosaur killer claws or climbing crampons?. In: Biology Letters. 2, Nr. 1, 2005, S. 110–112, doi:10.1098/rsbl.2005.0395.
- ↑ Phillip L. Manning u. a.: Biomecanics of Dromaeosaurid Dinosaur Claws: Application of X-Ray Microtomography, Nanoindentation, and Finite Element Analysis. In: The Anatomical Record. Advances in Integrative Anatomy and Evolutionary Biology. 292, Nr. 9, 2009, S. 1397–1405].
- ↑ W. Desmond Maxwell, John H. Ostrom: Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations. In: Journal of Vertebrate Paleontology. 15, Nr. 4, 1995, S. 707–712 (Zusammenfassung).
- ↑ Brian T. Roach, Daniel L. Brinkman: A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs. In: Bulletin of the Peabody Museum of Natural History. 48, Nr. 1, 2007, S. 103–138, doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2.
- ↑ Philip J. Currie, Aase Roland Jacobsen: An azhdarchid pterosaur eaten by a velociraptorine theropod. In: Canadian Journal of Earth Sciences. 32, 1995, S. 922–925.
- ↑ Enpu Gong, Larry D. Martin, David A. Burnham, Amanda R. Falk: The birdlike raptor Sinornithosaurus was venomous. In: Proceedings of the National Academy of Sciences. 2009, doi:10.1073/pnas.0912360107.
- ↑ a b Xing Xu, Xiao-Lin Wang, Xiao-Chun Wu: A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China. In: Nature. 401, 1999, S. 262–266, doi:10.1038/45769.
- ↑ Xing Xu, Zhonghe Zhou, Xiaolin Wang: The smallest known non-avian theropod dinosaur. In: Nature. 408, 2000, S. 705–708, doi:10.1038/35047056.
- ↑ a b Ji Qiang, Mark A. Norell, Ke-Qin Gao, Shu-An Ji, Dong Ren: The distribution of integumentary structures in a feathered dinosaur. In: Nature. 410, 2001, S. 1084–1087, doi:10.1038/35074079.
- ↑ a b c d X. Xu, Z. Zhou, X. Wang, X. Kuang, F. Zhang, X. Du: Four-winged dinosaurs from China. In: Nature. 421, 2003, S. 335–340, doi:10.1038/nature01342.
- ↑ Alan H. Turner, Peter J. Makovicky, Mark A. Norell: Feather Quill Knobs in the Dinosaur Velociraptor. In: Science. 317, Nr. 5845, 2007, S. 1721, doi:10.1126/science.1145076.
- ↑ a b Luis Chiappe, Gareth Dyke: The early evolutionary history of birds. In: Journal of the Paleontological Society of Korea. 22, Nr. 1, 2006, S. 133–151].
- ↑ Philip J. Currie: Feathered dinosaurs and the origin of birds. In: A. Legakis, S. Sfenthourakis, R. Polymeni, M. Thessalou-Legaki (Hrsg.): The New Panorama of Animal Evolution. Proc. 18th Int. Congr. Zoology. Pensoft, Sofia 2003, S. 55–60.
- ↑ Sankar Chatterjee, R. Jack Templin: Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui. In: Proceedings of the National Academy of Sciences. 104, Nr. 5, 2007, S. 1576–1580, doi:10.1073/pnas.0609975104.
- ↑ a b S. A. Czerkas, D. Zhang, J. Li, Y. Li: Flying Dromaeosaurs. In: Sylvia J. Czerkas (Hrsg.): Feathered Dinosaurs and the Origin of Flight. The Dinosaur Museum, Blanding 2002, ISBN 1932075011, S. 16–26 (The Dinosaur Museum Journal. 1).
- ↑ a b Alan Feduccia, Theagarten Lingham-Soliar, J. Richard Hinchliffe: Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence. In: Journal of Morphology. 266, S. 125–166, doi:10.1002/jmor.10382].
- ↑ Mark A. Norell u. a.: A new dromaeosaurid theropod from Ukhaa Tolgod (Omnogov, Mongolia). In: American Museum Novitates. 3545, 2006, S. 1–51, doi:10.1206/0003-0082(2006)3545[1:ANDTFU]2.0.CO;2.
- ↑ a b c d Nicholas R. Longricha, Philip J. Currie: A microraptorine (Dinosauria–Dromaeosauridae) from the Late Cretaceous of North America. In: Proceedings of the National Academy of Sciences. 106, Nr. 13, 2009, S. 5002–5007, doi:10.1073/pnas.0811664106.
- ↑ Torsten van der Lubbe, Ute Richter, Nils Knötschke: Velociraptorine dromaeosaurid teeth from the Kimmeridgian (Late Jurassic) of Germany. In: Acta Palaeontologica Polonica. 54, Nr. 3, 2009, S. 401–408, doi:10.4202/app.2008.0007.
- ↑ S. J. Metcalf, R. F. Vaughan u. a.: A new Bathonian (Middle Jurassic) microvertebrate site, within the Chipping Norton Limestone Formation at Hornsleasow Quarry, Gloucestershire. In: Proceedings of the Geologists’ Association. 103, 1992, S. 321–342, doi:10.1016/S0016-7878(08)80129-0.
- ↑ S. H. Hwang, M. A. Norell, Q. Ji, K. Gao: New Specimens of Microraptor zhaoianus (Theropoda: Dromaeosauridae) from Northeastern China. In: American Museum Novitates. 3381, 2002.
- ↑ Larry D. Martin: A basal archosaurian origin for birds. In: Acta Zoologica Sinica. 6, Nr. 50, 2004, S. 978–990.
- ↑ A. H. Turner, S. H. Hwang, M. Norell: A small derived theropod from Öösh, Early Cretaceous, Baykhangor Mongolia. In: American Museum Novitates. Nr. 3557, 2007, S. 1–27, doi:10.1206/0003-0082(2007)3557[1:ASDTFS]2.0.CO;2.
- ↑ X. Zheng, X. Xu, H. You, Q. Zhao, Z. Dong: A short-armed dromaeosaurid from the Jehol Group of China with implications for early dromaeosaurid evolution. In: Proceedings of the Royal Society B. 277, Nr. 1679, 2010, S. 211–217, doi:10.1098/rspb.2009.1178.
- ↑ Xing Xu u. a.: A new dromaeosaurid (Dinosauria: Theropoda) from the Upper Cretaceous Wulansuhai Formation of Inner Mongolia, China. In: Zootaxa. Nr. 2403, 2010, S. 1–9.
- ↑ Mark Norell, Peter Makovicky: Dromaeosauridae. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2. Auflage. University of California Press, Berkeley 2004, ISBN 0-520-24209-2, S. 196–209.
- ↑ William D. Matthew, Barnum Brown: The family Deinodontidae, with notice of a new genus from the Cretaceous of Alberta. In: Bulletin of the American Museum of Natural History. 46, 1922, S. 367–385.
- ↑ Ben Creisler: Dinosauria Translation and Pronunciation Guide. Abgerufen am 26. September 2009 (englisch).
- ↑ Henry F. Osborn u. a.: Three new Theropoda, Protoceratops zone, central Mongolia. In: American Museum Novitates. 144, 1924, S. 1–12.
- ↑ John H. Ostrom: Archaeopteryx and the origin of birds. In: Biological Journal of the Linnean Society. 8, 1976, S. 91–182, doi:10.1111/j.1095-8312.1976.tb00244.x.
- ↑ Zhonghe Zhou, Julia A. Clarke, Fucheng Zhang: Archaeoraptor’s better half. In: Nature. 420, Nr. 21, 2002, S. 285 (Abstract).
- ↑ Robert T. Bakker: Raptor Red. Bantam, New York 1995, ISBN 0-553-57561-9, S. 4.
Literatur
- Mark Norell, Peter Makovicky: Dromaeosauridae. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2. Auflage. University of California Press, Berkeley 2004, ISBN 0-520-24209-2, S. 196–209.
Weblinks
 Commons: Dromaeosauridae – Sammlung von Bildern, Videos und Audiodateien
Commons: Dromaeosauridae – Sammlung von Bildern, Videos und Audiodateien

Dieser Artikel wurde am 3. Dezember 2009 in dieser Version in die Liste der exzellenten Artikel aufgenommen. Kategorien:- Wikipedia:Exzellent
- Dromaeosauridae
- Dromaeosauriden
- Dromaeosauridae












Wikimedia Foundation.