- Cochlea
-
 Ausguss eines menschlichen Labyrinths
Ausguss eines menschlichen LabyrinthsDie Hörschnecke (lat. Cochlea) ist ein Teil des Innenohrs und stellt das Rezeptorfeld für die Hörwahrnehmung dar.
Inhaltsverzeichnis
Aufbau der Hörschnecke
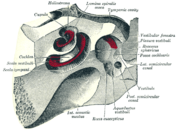 Aufbau der Hörschnecke
Aufbau der Hörschnecke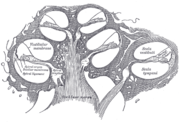 Querschnitt durch die Hörschnecke
Querschnitt durch die HörschneckeDie Hörschnecke ist ein schneckenförmiger Hohlraum im Felsenbein. Sie ist von einem Knochenmaterial umgeben, das nach den Zähnen das härteste Material im menschlichen Körper darstellt.
Die knöcherne Achse der Cochlea wird Modiolus (Schneckenspindel) genannt. Er steht mit dem inneren Gehörgang in Verbindung und enthält den Stamm des Höranteils des Nervus vestibulocochlearis (VIII. Hirnnerv).
Innen ist die Hörschnecke in drei übereinander liegende flüssigkeitsgefüllte Gänge gegliedert. Sie heißen
- Scala vestibuli (Vorhoftreppe; Scala, lat.: Treppe, Leiter; Vestibulum, lat.: Vorhof)
- Scala media oder Ductus cochlearis (Schneckengang) und
- Scala tympani (Paukentreppe).
Die Basis der Hörschnecke grenzt an das Mittelohr mit den Gehörknöchelchen. Die Fußplatte des Steigbügels ist in das ovale Fenster (Fenestra vestibuli oder Fenestra ovalis) beweglich eingepasst. Hinter dem ovalen Fenster liegt die Scala vestibuli. Diese ist an der Spitze der Schnecke (lat. Apex) über das Helicotrema (griechisch: Schneckenloch) mit der Scala tympani verbunden. Letztere grenzt an der Basis an das runde Fenster (Fenestra cochleae), welches durch das frei schwingende sekundäre Trommelfell (Membrana tympani secundaria) zum Mittelohr hin verschlossen ist. Ein Druck der Gehörknöchelchen auf das ovale Fenster läuft als Wanderwelle über die Scala vestibuli in Richtung zur Spitze der Schnecke und führt zur Auslenkung der Basilarmembran, was den eingetragenen Druck auf die Scala tympani überträgt. Über das runde Fenster kann dieser Druck ausgeglichen werden.
Die Scala media ist durch die Reißner-Membran (nach Ernst Reißner) von der Scala vestibuli und durch die Basilarmembran (Lamina basilaris oder Membrana basilaris) von der Scala tympani getrennt. Die Scala vestibuli und die Scala tympani sind mit Perilymphe gefüllt, die sich über das Helicotrema zwischen beiden Gängen austauscht. Die Scala media enthält Endolymphe. Beide Flüssigkeiten unterscheiden sich in ihrer Zusammensetzung: Die Perilymphe ähnelt dem extrazellulären Milieu, während die Endolymphe eine hohe Kalium-Konzentration aufweist. Darin gleicht sie dem Zytosol.
Vom Schall zum Nervenimpuls
 Schnitt durch die Hörschnecke: Aufbau des Corti-Organs
Schnitt durch die Hörschnecke: Aufbau des Corti-OrgansAuf der Basilarmembran liegt das Corti-Organ mit vier Reihen von Haarzellen, die unterschiedliche Eigenschaften haben:
- Die äußeren Haarzellen (drei Reihen) dienen der Verstärkung der Schallwanderwellen innerhalb der Cochlea (sog. Cochleärer Verstärker). Sie arbeiten als akustisches Vorfilter, das Dank der Fähigkeit der äußeren Haarzellen zur direkt vom Frequenzspektrum gesteuerten Längenverstellung auch ein schnell adaptives Filter ist.
- Die inneren Haarzellen (eine Reihe) leisten die Umwandlung mechanischer Schwingungen in Nervenimpulse (die so genannte Transduktion), die an das Gehirn weitergeleitet werden.
Die Umsetzung von Schall in Nervenimpulse hängt im Wesentlichen von folgenden Einflüssen ab:
- Elektrische und mechanische Schwingungseigenschaften von Zellkörper und Haarbündel der äußeren Haarzellen mit stetigen Gradienten entlang des Schneckenganges
- Umsetzung der mechanischen Anregung der inneren Haarzellen in Nervenimpulse
Schwingungsmechanische Eigenschaften des Innenohres
Tritt der Schall in das Innenohr ein, erzeugt er dort eine Welle, die durch das Innenohr wandert. Man spricht von der Wanderwelle. Sie lenkt mittels der Tektorialmembran die Sinneshärchen der äußeren, nicht jedoch die der inneren Haarzellen aus (Die äußeren Haarzellen, nicht jedoch die inneren, berühren über der Basilarmembran eine zweite Membran, die so genannte Tektorialmembran).
Basilarmembran und Schneckenkanal wirken hierbei als mechanisches Resonatorsystem. Da die Breite der Basilarmembran und der Durchmesser des Schneckenkanals vom ovalen Fenster zum Helicotrema hin zunehmen, ändern sich die mechanischen Eigenschaften (Massenbelag, Steife, Dämpfung) und damit auch die Schwingungseigenschaften des Systems in Abhängigkeit vom Abstand zum Helicotrema. Dies führt dazu, dass die Basilarmembran für unterschiedliche Frequenzen an unterschiedlichen Stellen in Resonanz gerät. Durch die hohe Steife der Basilarmembran erzeugen hohe Frequenzen in der Nähe des ovalen Fensters ein Auslenkungsmaximum, tiefe Frequenzen dagegen erst in der Nähe des Helicotrema. Für diese Erkenntnisse erhielt Georg von Békésy 1961 den Nobelpreis.
Die äußeren Haarzellen
Die äußeren Haarzellen reagieren bereits auf eine geringe Auslenkung ihrer Haarbündel mit einer aktiven Längenänderung ihres gesamten Zellkörpers. Dazu sind die äußeren Haarzellen durch ein besonderes Membranprotein, das Prestin befähigt. Das ist ein kontraktiles Protein in der Plasmamembran, das sich potenzialabhängig verkürzt oder verlängert. Transgene Mäuse, denen das Gen für Prestin fehlt, haben eine stark herabgesetzte Hörempfindlichkeit. Dies gilt als Beweis dafür, dass die Prestin-Motoren in der Zellmembran der äußeren Haarzellen den Schall innerhalb des Innenohres verstärken und die Frequenzselektivität erhöhen.
Die äußeren Haarzellen beeinflussen die mechanischen Schwingungen des Systems Basilarmembran-Schneckenkanal. An der Resonanzstelle werden die Schwingungen verstärkt und hierdurch die inneren Haarzellen stärker stimuliert. Jenseits der Resonanzstelle werden die Schwingungen stark gedämpft, die entsprechende Frequenz breitet sich kaum weiter aus. Hierdurch wird die Frequenzselektivität des Innenohres größer, die Zerlegung von Klängen oder von menschlicher Sprache in einzelne Tonfrequenzen wird erleichtert („cochleärer Verstärker“).
Ein weiterer Effekt ist, dass hohe Frequenzen, die in der Nähe des ovalen Fensters ihre Resonanzstelle besitzen, keine Anregung der inneren Haarzellen für tiefe Frequenzen bewirken. Tiefe Frequenzen, die erst in der Nähe des Heliocotremas ein Erregungsmaximum hervorrufen, erregen andererseits aber auch die für hohen Frequenzen zuständigen Haarzellen.
Die inneren Haarzellen
Die einzelnen Frequenzen eines auf diese Weise zerlegten Klangs reizen die auf die jeweiligen Frequenzen spezialisierten inneren Haarzellen. Der Reiz löst ein elektrisches Signal in den Haarzellen aus (mechano-elektrische Transduktion). Diese geben ein chemisches Signal (Transmitter Glutamat) an eine Hörnervenfaser (Transformation), wobei jede Hörnervenfaser wieder für eine einzelne Frequenz zuständig ist. Die Hörnervenfasern reagieren elektrisch (Aktionspotential) und reichen bis zum Stammhirn. Auf diese Weise werden die Tonfrequenzen getrennt und elektrisch zum Gehirn gesendet.
Die Erregung einer Haarzelle ist abhängig von der Vorgeschichte. Tritt eine mechanische Anregung nach einer gewissen Ruhephase ein, so „feuert“ die Haarzelle besonders intensiv. Bleibt die Anregung eine gewisse Zeit bestehen, so nimmt die Anzahl der Nervenimpulse ab (so genannte Adaptation). Erst nach einer gewissen anregungsarmen Zeit wird die ursprüngliche hohe Nervenimpulszahl wieder erreicht. (Dieser Sachverhalt wird unter anderem in psychoakustisches Modellen mit digitalen Signal-Prozessoren nachgebildet, die für die Audiodatenkompression in der Tonaufzeichnung verwendet werden.)
Innervation der Haarzellen
Die Haarzellen werden von afferenten und efferenten Nervenfasern versorgt. Während die afferenten Fasern aus dem Ganglion spirale stammen, kommen die efferenten Fasern aus den Olivenkernen (Tractus olivocochlearis oder Rasmussen-Bündel).
Das Ganglion spirale besteht aus über 30.000 bipolaren Nervenzellen. Mehr als 90 % davon sind myelinisierte Neurone (Typ I), die mit den inneren Haarzellen in Kontakt stehen. Die kleineren, unmyelinisierten Neurone (Typ II) versorgen die äußeren Haarzellen. Beide Typen senden Impulse an die Cochleariskerne in der Medulla oblongata.
Die efferenten Fasern laufen zuerst mit dem Nervus vestibularis in den inneren Gehörgang, zweigen dann jedoch über die Oort'sche Anastomose zum Cochlearisteil des Nervus vestibulocochlearis ab.
Die inneren Haarzellen werden von den radialen afferenten Fasern und den lateralen efferenten Fasern versorgt, die äußeren Haarzellen von den spiraligen afferenten und den medialen efferenten Fasern.
Innere Haarzellen: Alle aus dem Ganglion spirale stammenden Typ-I-Nervenzellen sind nur mit den inneren Haarzellen mit Synapsen verbunden. Ihre Dendriten bilden das radiale afferente System. Die Axone sammeln sich im Modiolus und verlaufen mit dem Nervus cochlearis zu den Cochleariskernen. Jede innere Haarzelle hat etwa mit zehn afferenten Fasern Kontakt, diese Zahl ist jedoch im Bereich des besten Hörens deutlich höher. Als Neurotransmitter für diese Synapsen wird Glutamat angenommen.
Ein Teil der efferenten Fasern (laterales efferentes System) erlangt Kontakt zu den Synapsenköpfchen der afferenten Fasern an den inneren Haarzellen und bildet mit ihnen Synapsen, diese Fasern haben also keinen direkten Kontakt mit Haarzellen.
Äußere Haarzellen: Die äußeren Haarzellen haben eine nur vergleichsweise geringe Versorgung mit afferenten Fasern. Nur in der apikalen Windung besteht eine höhere Versorgung mit afferenten Fasern. Die äußeren Haarzellen bilden ausschließlich Synapsen zu den unmyelinisierten Typ-II-Fasern des Ganglion spirale. Diese Fasern verlaufen am Boden des Corti-Tunnels von den äußeren Haarzellen in Richtung Modiolus, sie begleiten die äußeren Haarzellen spiralig (spiralig afferentes System), und jede einzelne Faser hat zu mehreren Zellen synaptischen Kontakt.
Die medialen efferenten Fasern ziehen als radiäre Tunnelfasern frei durch den Corti-Tunnel und bilden mit den Basen der äußeren Haarzellen Synapsen.
Einflüsse auf die akustische Wahrnehmung
Die Art und Weise, wie Schallsignale in Nervenimpulse umgesetzt werden, und an welcher Stelle Nervenimpulse im Innenohr entstehen, beeinflusst die akustische Wahrnehmung.
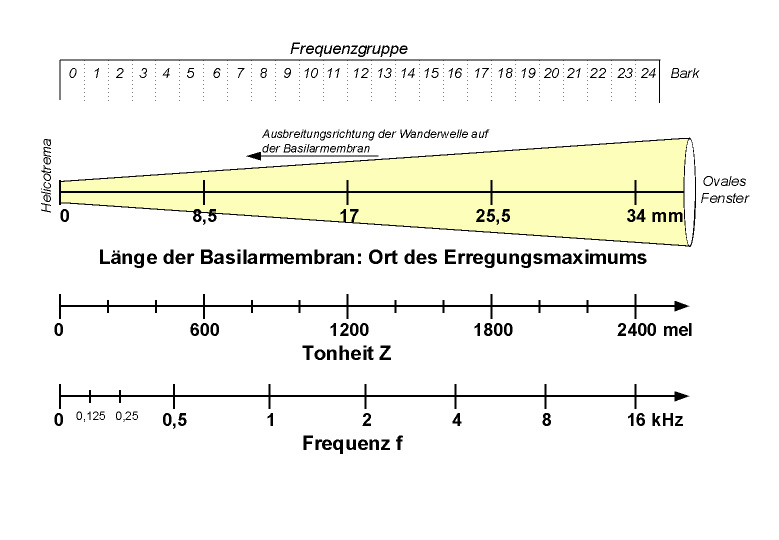
 Zusammenhang zwischen Basilarmembranort, Tonheit in Mel und Frequenz eines Tons
Zusammenhang zwischen Basilarmembranort, Tonheit in Mel und Frequenz eines TonsTonhöhe
Die Tonhöhe, die bei Beschallung mit einer gewissen Frequenz wahrgenommen wird, hängt eng mit dem Ort auf der Basilarmembran zusammen, an dem bei dieser Frequenz ein Erregungsmaximum besteht. Da die Basilarmembran am ovalen Fenster schmal und dick ist, ist ihre Eigenfrequenz hier hoch (bei geringer Amplitude). Weiter in Richtung Helicotrema, wo sie breiter und dünner ist, schwingt sie mit niedrigerer Frequenz (bei größerer Amplitude).
In Tierversuchen konnte man den Ort auf der Basilarmembran bestimmen, bei dem für eine bestimmte Frequenz eine maximale Erregung der inneren Haarzellen erzielt wird. Hieraus wurde anhand von physiologischen Vergleichen der Ort des Erregungsmaximums beim Menschen abgeleitet. Mit Hilfe von Hörversuchen kann wiederum bestimmt werden, welche Frequenz zu welcher Tonhöhenempfindung führt. Man fand einen linearen Zusammenhang zwischen der Position des Erregungsmaximums auf der Basilarmembran (gerechnet als Abstand vom Helicotrema) und der empfundenen Tonhöhe.
Das menschliche Ohr ist im Stande, Frequenzunterschiede von nur 3 Hz zu detektieren; der Hörbereich geht etwa von 20 Hz bis 20 kHz, wobei die Hörschwelle für verschiedene Frequenzen unterschiedlich ist und bei 2-4 kHz am niedrigsten ist; biographisch hört man vor allem die hohen Frequenzen zunehmend schlechter (Presbyakusis). Ortsprinzip: Wird die Basilarmembran mit einer bestimmten Frequenz angeregt, so wird sie dort am meisten schwingen, wo sie am besten mit dieser Frequenz schwingen kann. Für eine Frequenz sind daher nur wenige innere Haarzellen verantwortlich (die für diese Frequenz auch eine besonders niedrige Reizschwelle haben); aufgrund der Verschaltung in der Hörbahn sind auch im primären auditiven Cortex bestimmte Neurone nur für bestimmte Frequenzen zuständig (Tonotopie. Dieses Schwingungsmaximum der Basilarmembran ist neben dieser passiven Komponente vor allem auch deshalb scharf umrissen, weil die Äußeren Haarzellen am Ort des Schwingungsmaximums erregt werden und die Schwingung durch Kontraktion etwa tausendfach verstärken (cochleärer Verstärker), so dass nur in einem sehr sehr kleinen Gebiet die Inneren Haarzellen sehr stark erregt werden. Zu dieser „mechanisch aktiven“ Komponente kommt noch eine neuronale: laterale Hemmung entlang der Hörbahn (vor allem im Ganglion spirale cochleae), d. h. stark erregte Neurone hemmen benachbarte leicht erregte Neurone (die den direkt benachbarten Frequenzbereich übertragen würden). Diese Kontrastierung dient der Rauschunterdrückung. Für Frequenzen über 4 kHz (die durchaus noch im Sprachbereich liegen) ergibt sich ein Problem daraus, dass die inneren Haarzellen „nur“ maximal etwa 4000-mal pro Sekunde feuern können, eine Schwingung von 4,2 kHz kann also nicht 1:1 an den N. cochlearis weitergegeben werden. Nun macht sich das menschliche Ohr aber das Prinzip der Phasenkodierung zu Nutze: Liegen zwischen zwei Sensorpotenzialen einer Haarzelle etwa einmal 6 ms, einmal 15 und einmal 9, so errechnet das Zentralnervensystem den größten gemeinsamen Teiler und erhält 3 ms als den Abstand der Wellenberge.
Lautstärke und Klang
Die Anzahl der insgesamt erzeugten Nervenimpulse per Frequenzband ist ein Maß für die empfundene Lautheitstärke eines Schallsignals. Die Anzahl der abgegebenen Nervenimpulse hängt wiederum von der Stärke der Erregung der inneren Haarzellen und damit vom Schwingungsverhalten der Basilarmembran ab.
 Maskierungeffekt als Indiz für das Erregungsverhalten des Innenohres
Maskierungeffekt als Indiz für das Erregungsverhalten des InnenohresDas Erregungsmuster, das ein bestimmter Ton verursacht, kann man anhand von Maskierungsexperimenten nachvollziehen. Ist bei Anwesenheit eines Tons ein zweiter leiserer Ton nicht mehr wahrnehmbar, so deutet dies darauf hin, dass der erste Ton die Nervenzellen, die für die Wahrnehmung des zweiten Ton verantwortlich sind, schon wesentlich stärker erregt hat, als es der zweite Ton kann.
Aufgrund des Schwingungsverhaltens der Basilarmembran regen Einzeltöne auch noch Nervenzellen an, die oberhalb ihrer Frequenz liegen, also zu Frequenzen gehören, die gar nicht im Schallsignal enthalten sind. Bei Schallsignalen mit flachem Frequenzgang werden keine Nervenzellen außerhalb des Frequenzgangs des Schallsignals erregt. Dies führt dazu, dass Einzeltöne (oder Schallsignale mit starken tonalen Anteilen) lauter empfunden werden, als breitbandige Schallsignale mit gleichem Schallpegel.
Auf der anderen Seite beeinflusst das Zeitverhalten eines Schallsignals die Anzahl der abgegebenen Nervenimpulse. Setzt (in einem Frequenzbereich) ein Schallsignal nach längerer Ruhe ein, feuern die Nervenzellen besonders stark. Bei länger andauerndem Schall sinkt die Anzahl der Nervenimpulse wieder auf einen durchschnittlichen Wert ab.
Dies führt dazu, dass Schallsignale mit plötzlichen Schalleinsätzen (z. B. Hämmern) als wesentlich lauter empfunden werden, als gleichförmige Schallsignale mit gleichem Schallpegel.
Ebenso wie die Lautstärke wird hierdurch auch der empfundene Klang beeinflusst, tonale Anteile und Schalleinsätze bestimmen so den Klangeindruck wesentlich stärker als es das physikalische Spektrum eines Schallsignals erwarten lässt.
Signalverarbeitung des Gehörs
Die Position des Erregungsmaximums auf der Basilarmembran bestimmt nicht nur die empfundene Tonhöhe (siehe oben), sondern auch, welche Signalanteile vom Gehör gemeinsam ausgewertet werden.
Hierzu teilt das Gehör die Basilarmembran in etwa 24 gleich lange Abschnitte ein, sogenannte Frequenzgruppen. Die Nervenimpulse aus einer Frequenzgruppe werden gemeinsam ausgewertet, um hieraus Lautstärke, Klang und Richtung des Schallsignals in diesem Frequenzbereich zu bestimmen.
Die Breite einer Frequenzgruppe beträgt ca. 100 Hz bei Frequenzen bis 500 Hz und eine kleine Terz oberhalb von 500 Hz. (Dies entspricht ca. 1 Bark oder 100 Mel)
Der gesunde Mensch kann normalerweise Frequenzen von 20-18000 Hz wahrnehmen. Der Frequenzumfang nimmt mit dem Alter ab.
Technische Anwendung von Innenohr-Effekten
Ausgenutzt werden Innenohr-Effekte bei Datenreduktionsverfahren wie MP3.
Hier werden, ebenso wie beim Gehör, Signalbereiche in Frequenzgruppen analysiert. Signalbereiche, die aufgrund von Maskierungseffekten (also aufgrund von Innenohrmechanik) nicht hörbar sind, werden aus dem Signal entfernt oder mit geringerer Qualität übertragen. Hierdurch sinkt die Datenmenge, ein Unterschied zum Ursprungssignal ist aber für den Menschen nicht wahrnehmbar.
Das gesunde Ohr weist ein Frequenzunterscheidungsvermögen bei 1 kHz von etwa +/- 3 Hz auf. Liegt eine Hörstörung vor, so kann das Frequenzunterscheidungsvermögen in Abhängigkeit von Art und Ausmaß der Hörstörung herabgesetzt sein.
Siehe auch
Literatur
- Martin Trepel: Neuroanatomie. Struktur und Funktion. StudentConsult. 3.. Auflage. Urban & Fischer Bei Elsevier, 2006, ISBN 3437444255.
- M. C. Liberman, J. Gao, D. Z. He, X. Wu, S. Jia, J. Zuo: Prestin is required for electromotility of the outer hair cell and for the cochlear amplifier.. In: Nature. 419, Nr. 6904, 2002, S. 300-304 (doi:10.1038/nature01059).
Weblinks





Wikimedia Foundation.