- Kabeltheorie
-
 Abb.1: Schematische Darstellung der Widerstände und Kapazitäten an einem Axon.
Abb.1: Schematische Darstellung der Widerstände und Kapazitäten an einem Axon.
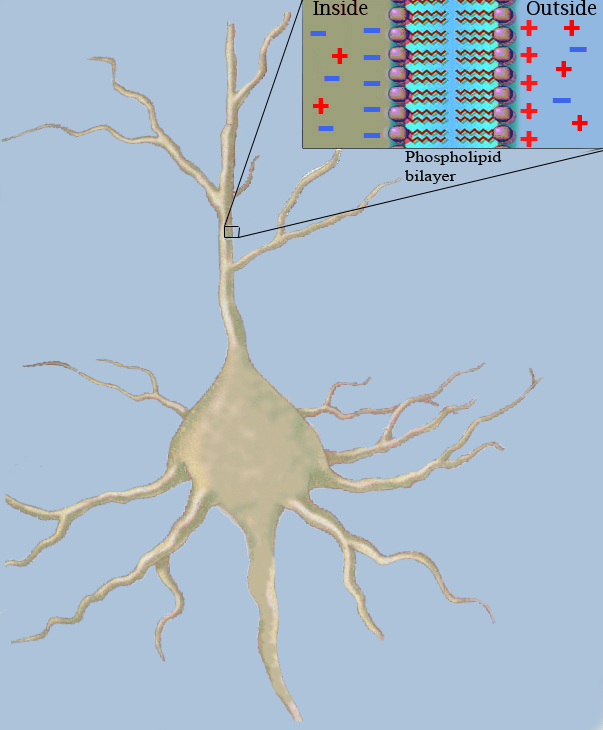
Die klassische Kabeltheorie versucht, die elektrischen Ströme und die Potentialdifferenzen entlang passiver Nervenfasern (Neuriten), insbesondere entlang von Dendriten, mit Hilfe von mathematischen Modellen zu beschreiben. Dazu werden die Nervenfasern vereinfachend als eine Aneinanderreihung von gleichartig aufgebauten zylindrischen Segmenten betrachtet. Die Wand jedes dieser Segmente wird durch die Lipiddoppelschicht der Axonmembran gebildet, deren elektrische Eigenschaften sich als die Parallelschaltung eines elektrischen Widerstandes rm und eines Kondensators mit der Kapazität cm beschreiben lassen. Die Kapazität cm der Nervenfaser resultiert aus den elektrostatischen Kräften, die aufgrund ungleicher Ladungsverteilung im Extra- und Intrazellulärraum über die sehr dünne Lipiddoppelschicht der Plasmamembran wirksam sind (siehe Abb. 2). In Längsrichtung der Nervenfaser besteht durch den Widerstand, den das Zytosol der Bewegung von elektrisch geladenen Teilchen entgegensetzt, der Längswidertand rl.
 Abb.2: Membrankapazität
Abb.2: MembrankapazitätInhaltsverzeichnis
Geschichte
Die Ürsprünge der Kabeltheorie reichen bis in die 50er Jahre des 19. Jahrhunderts zurück als William Thomson (später bekannt als Lord Kelvin) damit begann mathematische Modelle über den Signalabfall in telegrafischen Unterseekabeln zu entwickeln. Diese Modelle besaßen Ähnlichkeit mit den partiellen Differentialgleichungen, die von Fourier benutzt wurden um die Wärmeleitung in Festkörpern zu beschreiben.
Um 1870 war Hermann der Erste, der versuchte ein Modell des axonalen Elektrotonus' zu entwickeln, wobei er sich an den Analogien zur Wärmeleitung orientierte. Allerdings war es Hoorweg, der 1898 die Ähnlichkeit mit Kelvin's Unterseekabeln entdeckte. In der Folge waren es Hermann und Cremer, die unabhängig voneinander zu Beginn des 20. Jahrhunderts die Kabeltheorie für Nervenfasern entwickelten. Weitere mathematische Theorien über die Erregungsleitung in Nervenfasern, die auf der Kabeltheorie basierten, wurden von Cole und Hodgkin (1920-1930), Offner et al. (1940) und Rushton[1] [2] erarbeitet.
Experimentelle Belege für die Wichtigkeit der Kabeltheorie bei der Beschreibung von realen Nervenfasern wurden in den 30er Jahren durch Arbeiten von Cole, Curtis, Hodgkin, Bernard Katz, Rushton, Tasaki und anderen geliefert. Zwei wichtige Artikel dieser Zeit stammen von Davis und Lorente de No,[3] beziehungsweise Hodgkin und Rushton.[4]
Durch die Weiterentwicklung der Techniken zur Messung von elektrischer Aktivität an einzelnen Nervenfasern in den 50er Jahren erlangte die Kabeltheorie zunehmend Bedeutung bei der Analyse von mit Mikroelektroden intrazellulär abgeleiteten Strömen und Spannungen sowie bei der Untersuchung der elektrischen Eigenschaften neuronaler Dendriten. Wissenschaftler wie Coombs, Eccles, Fatt, Frank, Fuertos und Andere orientierten sich beim Design neuer Versuche zur Erlangung neuer Erkenntnisse der Funktionsweise von Neuronen nun an der Kabeltheorie.
Später erlaubte die Kabeltheorie mit ihren mathematischen Formeln immer ausgefeiltere Neuronenmodelle, die von Wisschenschaftlern wie Rall, Redman, Rinzel, Idan Segev, Tuckwell,[5] Bell, Poznanski and Ianella untersucht wurden.
Verschiedene wichtige Ansätze zur Erweiterung der Kabeltheorie haben in letzter Zeit zum Einsatz von Ionenkanälen und endogener Strukturen geführt, um die Effekte einer unterschiedlichen Verteilung des synaptischen Inputs über die dendritische Oberfläche zu untersuchen.[6]
Herleitung der Kabelgleichung
Die oben eingeführten Größen rm und cm werden in Abhängigkeit von der Faserlänge gemessen. Daher besitzt rm die Einheit [Ωm] und cm Farad pro Meter ([F/m]). Im Gegensatz dazu werden der spezifische Membranwiderstand Rm und die spezifische Membrankapazität Cm in Abhängigkeit von der Membranoberfläche gemessen und besitzen die Einheit [Ω m2] bzw. [F/m2]. Ist der Faserradius r, der vereinfachend als konstant angenommen wird, bekannt und der Faserumfang demnach
 , so können rm und cm wie folgt berrechnet werden:
, so können rm und cm wie folgt berrechnet werden: (1)
(1)
 (2)
(2)
Dass diese Gleichungen Sinn machen ist verständlich, wenn man sich klar macht, dass sich die Membranoberfläche proportional zum Faserumfang verhält: der Membranwiderstand nimmt mit zunehmendem Faserumfang ab, weil den geladenen Teilchen nun eine größere Fläche zum Durchtritt zur Verfügung steht. Entsprechend nimmt die Membrankapazität zu, weil durch den größeren Faserumfang, die Oberfläche des durch die Plasmamembran gebildeten Kondensators zunimmt.In ähnlicher Weise erlaubt der spezifische Widerstand Rl (Einheit: [Ωm]) des Zytoplasmas die Berechnung des intrazellulären Längswiderstandes rl der Faser mit der Einheit [Ω/m]:
 (3)
(3)
Um die Herleitung der Kabelgleichung besser zu verstehen, wird die einleitend beschriebene Nervenfaser zunächst noch weiter vereinfacht: es wird im Folgenden angenommen, dass sie eine für geladene Teilchen absolut undürchlässige Plasmamembran besitzt, dass also rm unendlich groß ist und daher kein Ladungsverlust nach außen auftritt. Ebenso sei die Membrankapazität null (cm = 0). Ein Strom, der unter diesen Annahmen, an Position x = 0 in die Nervenfaser injiziert wird, würde entlang dem Innern der Faser unverändert fortlaufen. Mit zunehmender Entfernung vom Ort der Injektion des Stroms kann mit Hilfe des Ohmschen Gesetzes die Änderung der Spannung berechnet werden:
 (4)
(4)
Lässt man Δ x gegen null gehen, so erhält man folgende partielle Differentialgleichung:
 (5)
(5)
oder
 (6)
(6)
Bezieht man nun die Tatsache, dass rm nicht unendlich groß ist wieder mit in die Überlegungen ein, ist dies als würde man Löcher in einen Gartenschlauch machen. Umso mehr Löcher der Schlauch enthält, desto mehr Wasser wird nach außen verloren gehen und desto weniger Wasser wird einen bestimmten Punkt des Schlauchs erreichen. In ähnlicher Weise wird in der Nervenfaser ein Teil des in Längsrichtung fließenden Stroms durch die Plasmamembran verloren gehen.Wenn im der Strom ist, der über die Membran pro Faserabschnitt verloren geht, dann ergibt sich die Summer aller Ströme die entlang von n Abschnitten verloren gehen zu
 . Die Stromänderung Δi im Zytoplasma nach Zurücklegen der Strecke Δx kann daher geschrieben werden als
. Die Stromänderung Δi im Zytoplasma nach Zurücklegen der Strecke Δx kann daher geschrieben werden als (7)
(7)
bzw.
 (8)
(8)
Unter Berücksichtigung, dass die Kapazität cm nicht null ist, kann im mit Hilfe einer weiteren Formel ausgedrückt werden. Die Kapazität cm erzeugt im Zytoplasma einen Ladungsfluss in Richtung zur Membran, der üblicherweise als Verschiebungsstrom bezeichnet wird und im Folgenden als ic geschrieben wird. Dieser Strom wird nur fließen solange die Membrankapazität ihren Endwert nicht erreicht hat. im kann dann ausgedrückt werden als: (9)
(9)
wobei
 die Potentialänderung in Abhängigkeit von der Zeit beschreibt. Der Strom ir, der über die Membran fließt, errechnet sich gemäß dem Ohmschen Gesetz wie folgt:
die Potentialänderung in Abhängigkeit von der Zeit beschreibt. Der Strom ir, der über die Membran fließt, errechnet sich gemäß dem Ohmschen Gesetz wie folgt: (10)
(10)
Weil weiter gilt, dass im = ir + ic ergibt sich für im: (11)
(11)
 gibt dabei die Änderung des in logitudinaler Richtung fließenden Stroms pro Faserabschnitt an.
gibt dabei die Änderung des in logitudinaler Richtung fließenden Stroms pro Faserabschnitt an.
Durch Einsetzen von Gleichung (6) in (11) erhalten wir als erste Version der Kabelgleichung folgende partielle Diffreentialgleichung zweiter Ordnung: (12)
(12)
Durch Umstellung von Gleichung (12) (siehe unten) erhält man zwei wichtige Größen, nämlich die Membranlängskonstante λ und die Membranzeitkonstante τ, die in den nächsten Abschnitten näher beschrieben werden.Membranlängskonstante
Die Membranlängskonstante λ mit der Einheit [m] ist ein Parameter, der angibt wie weit sich ein Strom entlang dem Innern der Nervenfaser ausbreiten und damit die Spannung entlang dieser Strecke beeinflussen wird. Je größer λ, desto weiter breitet sich der Strom aus. Aus dem Membranwiderstand rm und dem Längswiderstand rl errechnet sich die Längskonstante folgendermaßen:
 (13)
(13)
Diese Formel ist sinnvoll, denn je größer der Membranwiderstand rm, desto mehr Strom wird im Zytosol verbleiben um entlang der Nervenfaser zu wandern, was zu einer Vergrößerung der Membranlängskonstante führt. Gleiches gilt für eine Verringerung des Längswiderstandes rl, denn es ist nun einfacher für die Ladungsträger sich entlang der Nervenfaser zu bewegen. Löst man Gleichung (12), kommt man zu folgender Gleichung, die für einen stabilen Zustand gültig ist, zum Beispiel wenn die Zeit t gegen unendlich geht: (14)
(14)
V0 ist hierbei die Potentialdifferenz über der Plasmamembran an der Stelle x = 0 (Ort der Strominjektion), "e" die eulersche Zahl und Vx die Potentialdifferenz in der Entfernung "x" vom Ort der Strominjektion. Ist x = λ, dann gilt:
 (15)
(15)
und
 (16)
(16)
Wird die Potentialdifferenz V in der Entfernung x = λ von x = 0 gemessen, erhält man
 (17)
(17)
Daraus folgt, dass Vλ immer 36,8 Prozent von V0 beträgt. Die Membranlängskonstante gibt folglich genau jene Strecke in Metern an, an der die Potentialdifferenz auf 36,8 Prozent von V0 abgefallen ist.
Membranzeitkonstante
Neurobiologen sind häufig daran interessiert zu wissen wie schnell sich das Membranpotential Vm einer Nervenfaser, als Reaktion auf eine Änderung des Stroms der in das Zytosol injiziert wird, ändert und beschreiben dazu die Membranzeitkonstante τ. Sie gibt die Zeit in Sekunden an, nach der die Amplitude der Potentialdifferenz auf 36,8% des Ausgangswertes abgesunken ist. Sie ist damit ein Maß für die Geschwindigkeit der Potentialänderung und lässt sich wie folgt errechnen:
 (18)
(18)
Kabelgleichung mit Membranlängs- und zeitkonstante
Multipliziert man Gleichung (12) auf beiden Seiten mit rm ergibt sich:
 (19)
(19)
Ersetzt man nun
 durch λ2 und
durch λ2 und  durch τ erhält man die vielleicht bekannteste Form der Kabelgleichung:
durch τ erhält man die vielleicht bekannteste Form der Kabelgleichung: (20)
(20)
Anmerkungen
Einleitend wird erwähnt, dass die Kabeltheorie die Verhältnisse an passiven Nervenfasern beschreibt. Passiv meint in diesem Fall die Unabhängigkeit des Membranwiderstands rm vom Membranpotential. Neuere Untersuchungen mit dendritischen Membranen haben allerdings gezeigt, dass viele von ihnen mit spannungsabhängigen Ionenkanälen ausgestattet sind und der Membranwiderstand daher sehr wohl vom Membranpotential abhängt.[7] Folglich ist eine Anpassung der klassischen Kabeltheorie erforderlich um dieser Tatsache Rechnung zu tragen.
Einzelnachweise
- ↑ Lussier JJ, W. A. H. Rushton: The relation between the space constant and conduction velocity in nerve fibers of the A group from the frog's sciatic. In: The Journal of Physiology. 114, Juli 1951, S. 399-409. PMID 14861824.
- ↑ W. A. H. Rushton: A theory of the effects of fibre size in medullated nerve. In: The Journal of Physiology. 115, September 1951, S. 101-122. PMID 14889433.
- ↑ DAVIS L Jr, LORENTE de NO R.: Contribution to the mathematical theory of the electrotonus. In: Rockefeller Institute for Medical Research. 131, 1947, S. 442-496. PMID 20261883.
- ↑ A. L. Hodgkin, W. A. H. Rushton: The electrical constants of a crustacean nerve fibre. In: Proceedings of the Royal Society of Medicine. 133, Nr. 873, Dezember 1946, S. 444-479. PMID 20281590.
- ↑ Henry C. Tuckwell: Introduction to Theoretical Neurobiology. Cambridge University Press 4 1988, ISBN 0521350964
- ↑ Roman R. Poznanski: Thermal noise due to surface-charge effects within the Debye layer of endogenous structures in dendrites. In: Physical Review. 81, Nr. 2, Februar 2010. PMID 20365590.
- ↑ Greg J. Stuart, Bert Sakmann: Active propagation of somatic action potentials into neocortical pyramidal cell dendrites. In: Nature. 367, Januar 1994, S. 69-72. PMID 8107777.

Wikimedia Foundation.